
ESTIMATION OF ENVIRONMENTAL CAPACITY OF HUCHO BLEEKERI IN TAIBAI RIVER BASED ON ECOPATH MODEL
-
摘要:
为评估太白河川陕哲罗鲑(Hucho bleekeri Kimura)的资源现状和环境容纳量, 于2017—2018年对太白河的鱼类资源、饵料生物资源、水体理化特征等进行了调查。采用密度法估算出太白河川陕哲罗鲑现存资源量为3814尾, 95%的置信区间为3624—4005尾; 构建了太白河流域的Ecopath模型, 河流总生产量为1748.91 t/(km2·a), 其中初级生产力为1643.00 t/(km2·a), 生态系统的连接指数(CI)和系统杂食指数(SOI)分别为0.429和0.087。川陕哲罗鲑、秦岭细鳞鲑(Brachymystax lenok tsinlingensis)、小型中下层鱼类和浮游植物四个功能组为太白河水域生态系统中的关键种(类群), 估算出川陕哲罗鲑的环境容纳量为1.87 t/km2, 约5127尾, 其中幼鱼为3399尾, 成鱼为1728尾。基于现存量和环境容纳量, 建议强化栖息地保护, 促进川陕哲罗鲑资源恢复。
Abstract:Hucho bleekeri, a distinctive and regionally distributed cold-water fish in China, attained the status of a national first-class protected animal in 2021. To explore the current population of Hucho bleekeri in the Taibai River Basin, we estimated the existing resources based on biological survey data from the Taibai River conducted across four seasons from 2017 to 2018, utilizing systematic sampling techniques and the river reach method. Employing EwE software, we constructed an Ecopath model to depict the historical habitat of Hucho bleekeri in the Taibai River basin. The results showed that the total amount of Hucho bleekeri in the Taibaihe River was 3814 fish, with a 95% confidence interval ranging from 3624 to 4005 fish. Within this, the adult fish resources amounted to 1286, with a 95% confidence interval from 1233 to 1339, and juvenile fish resources accounted for 2528, with a 95% confidence interval ranging from 2438 to 2619. The carrying capacity of historical habitat environment for Hucho bleekeri in the Taibai River basin was 1.87 t/km2, supporting approximately 5127 dult fish and 3399 young fish per kilometer square, with an additional 1728 fish for tail areas. The total productivity of the ecosystem was 1748.91 t/(km2·a), with total primary productivity at 1643.00 t/(km2·a), constituting 93.94% of the total productivity. The aquatic ecosystem displayed a connectivity index (CI) of 0.429 and s system omnivore index (SOI) of 0.087, respectively. Zooplankton exhibited the highest ecological nutrient conversion efficiency at 0.84. Key species identified within the ecosystem included Hucho bleekeri, Brachymystax lenok tsinlingensis, small middle and lower strata fishes. and phytoplankton. In conclusion, the existing resources of Hucho bleekeri in their historical habitats in the Taibai River Basin fall below their environmental capacity. Therefore, it is recommended that to enhance investigations of spawning grounds and habitat protection to facilitate the recovery of Hucho bleekeri.
-
Keywords:
- Ecopath model /
- Environmental accommodation /
- Taibai River /
- Hucho bleekeri
-
川陕哲罗鲑(Hucho bleekeri Kimura)又称虎嘉鱼、贝氏哲罗鲑、布氏哲罗鲑、四川哲罗鲑, 隶属于鲑形目(Salmoniformes)鲑科(Salmonidae)哲罗鲑属(Hucho), 是我国特有的、局部分布的冷水性鱼类。川陕哲罗鲑分布于四川省岷江上游(最初分布的下限是灌县)、大渡河上游(最初分布下限是峨边)、陕西的太白河及留坝的汉水上游、青海的马柯河[1, 2]。在20世纪60、70年代后, 由于拦河筑坝、兴建水利工程、挖沙、开采金矿及电鱼毒鱼等人类活动, 致使川陕哲罗鲑的生态环境遭到破坏, 分布区域不断缩小, 川陕哲罗鲑资源量开始大幅度减少[3]。到20世纪80年代, 川陕哲罗鲑分布范围缩减, 其分布区域为岷江上游、大渡河上游和青衣江上游。至20世纪末, 川陕哲罗鲑野外资源量极其稀少, 仅在大渡河上游、岷江上游的黑水河及陕西省汉江支流的太白河有少量发现。1988年国务院将其列为国家二级保护动物, 1998年列为濒危物种[4], 2021年升级为国家一级保护动物。
太白河位于秦岭南麓, 发源于鳌山南麓。主流经嘴头镇南部, 贯穿太白河乡, 于元坝子入留坝县江口镇汇入太白河。沿途纳苏家沟、无名沟、耳爬沟、双合沟、王家沟、庙沟、东沟好桑园坝河等小型支流。在太白县集流面积377.87 km2, 发源高程海拔2470 m, 出境高程海拔1100 m, 平均坡降30.9‰, 平均流量5.2 m3/s, 森林覆盖率达89.5%。2012年9月, 在陕西省太白河洪水期后, 获得了19尾川陕哲罗鲑成体, 这是绝迹15年后的再次发现, 也是近30多年来在全国范围内发现的规模最大的一批[5]。目前, 川陕哲罗鲑的研究主要是集中在生物学、摄食、繁殖、栖息地环境和资源调查等方面, 而对川陕哲罗鲑栖息地特征和资源量评估的研究较少。太白河作为目前川陕哲罗鲑主要栖息地, 对太白河川陕哲罗鲑种群进行研究具有重要保护意义。
Ecopath模型被国际上许多生态学家认为是新一代水域生态系统研究的核心工具, 用于定量分析生态系统营养结构、不同功能组间能量流动特点及科学评估人类活动对生态系统的影响[6]。Ecopath模型主要用于研究食物网中物种间的捕食、竞争关系及对物种生物量的估算[7, 8]。近年来, 该模型作为评估增殖放流活动中物种环境容纳量的工具被广泛关注[9, 10]。在濒危水生动物保护方面, 吴斌等[10]利用Ecopath模型估算了鄱阳湖通长江水道长江江豚的环境容纳量, 为鄱阳湖长江江豚的保护和科学管理提供了理论依据。因此, 本研究在实地调查的基础上通过Ecopath with Ecosim软件构建太白河水生态系统食物网模型, 系统分析生态系统特征和能量流动格局, 估算川陕哲罗鲑的环境容纳量。同时采用密度法, 估算太白河川陕哲罗鲑的现存资源量, 分析太白河川陕哲罗鲑种群衰退的原因, 为太白河川陕哲罗鲑种群恢复工作提供科学支撑。
1. 材料与方法
1.1 采样点设置
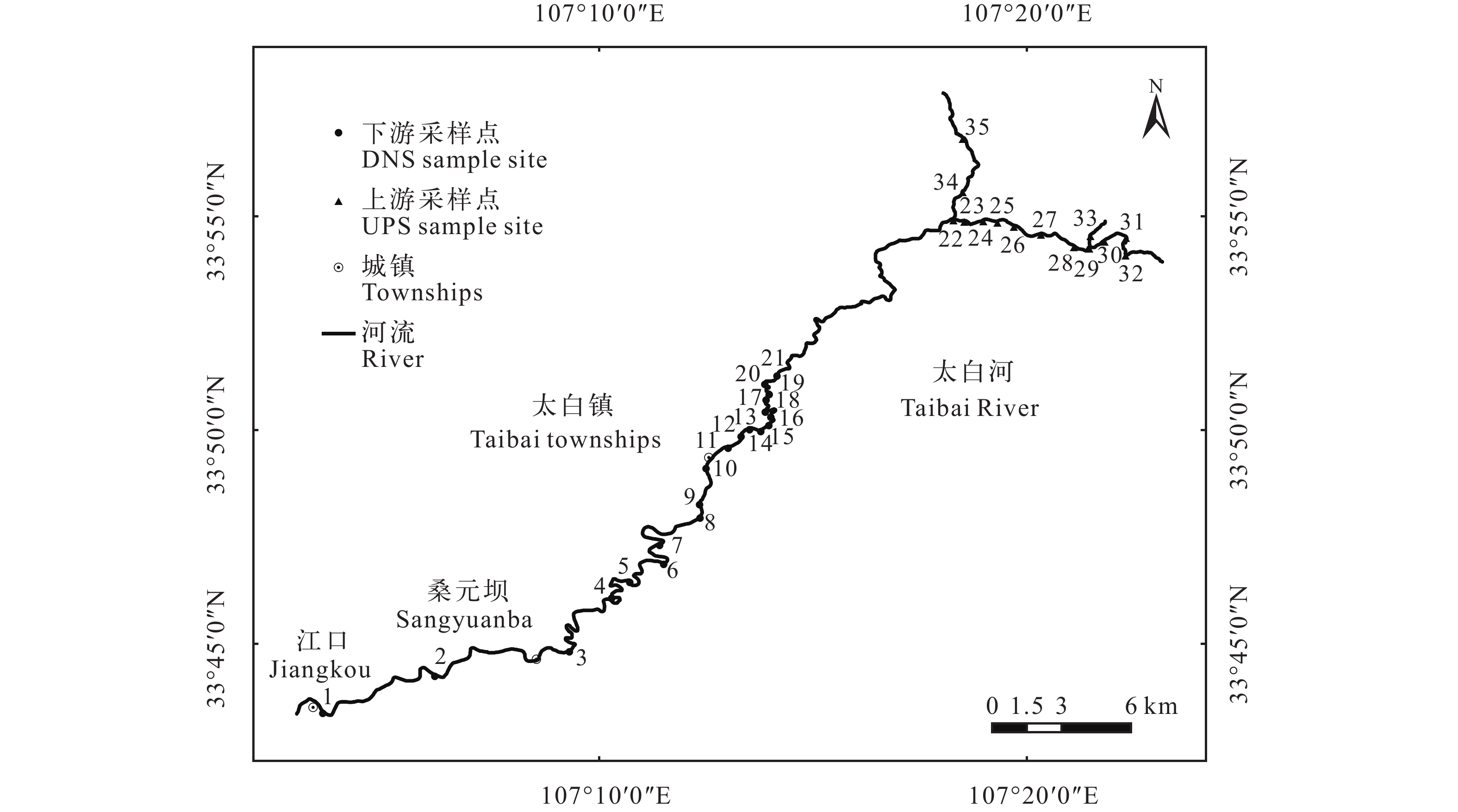
根据川陕哲罗鲑的历史分布, 在太白河上下游设置35个采样点(其中21#和22#样点之间的河段为管制区域, 无法进入采集; 图 1)。
1.2 调查方法
2017—2018年, 对太白河流域进行文献调研和实地调查。流域特性主要通过文献调研法收集, 水体理化特性和水生生物现状调查分别于2017年10月、2018年1月, 2018年4月和2018年7月共开展四次(表 1)。其中, 鱼类调查仅在2017年10月开展1次, 具体方法: 在获得地方主管部分许可后, 采用电捕法按照从下游到上游的路径进行, 1人背负电鱼机(4500 W)持续放电, 1人持抄网捞取电晕的鱼类样本, 每个样点捕捞50—100 m长度, 每个样点重复捕捞2次。为防止鱼类逃逸, 在调查样点的上下游设置网目尺寸为2 cm的网, 防止鱼类逃逸。捕获的鱼类样本经鉴定、测量、记录后原地放流。
表 1 太白河基本环境特征调查内容Table 1. Basic environmental characteristics of the Taibai River survey content类别
Category调查内容
Investigation content断面及频次
Section and frequency数据来源
Data source调查方法
Investigation method流域特性
Characteristics of river basin流量、流域面积、比降、植被覆盖、人口 整体1次 文献 通过查阅地方志、统计年鉴等资料获得 水体理化特性
Physicochemical characteristics水温、溶氧、流速、透明度、pH 在A段和C段分别设置1个和2个断面, 按照季节调查4次 实地测量 采用美国YSI公司 Proplus 便携式水质监测仪进行测量 水生生物现状
Aquatic life status鱼类、浮游动植物、底栖生物、水生高等植物 与水体理化特性相同 实地采集 参考《河流水生态调查技术方法》[11] 1.3 川陕哲罗鲑资源量计算方法
(1)各样点川陕哲罗鲑的密度($ {D}_{i} $)
$$ {D}_{i} = \frac{{N}_{i}}{{L}_{i}} $$ 式中, $ {D}_{i} $为每一个样点的鱼类密度(ind./m); $ {N}_{i} $为每一样点采集的川陕哲罗鲑数目(ind.); $ {L}_{i} $为每一个样点捕捞的河段距离长度。
(2)川陕哲罗鲑种群的平均出现密度($\bar{D}$)
$$ \bar{D} = \frac{\sum _{i=1}^{n}{D}_{i}}{n} $$ 式中, n为采集到川陕哲罗鲑的样点数。
抽样绝对误差($ \text{∆} $)
$$ \text{∆} = {t}{\alpha }\cdot {\mu }_{x} $$ 取 $ \alpha $=95%水平时, $\text{∆}$=1.96$\cdot {\mu }_{x}$, 其中 $ {\mu }_{x} $为抽样调查标准误, 计算公式:
$$ {\mu }_{x} = \frac{\sigma }{\sqrt{n}} $$ $$ \sigma = \frac{\sum _{i=1}^{n}{{(D}_{i} - \bar{D})}^{2}}{n - 1} $$ 估算出川陕哲罗鲑资源量后, 结合渔获物组成情况估算出其他鱼类的资源量。
1.4 模型构建方法
采用EwE (版本6.6.6.17608)软件进行建模和分析。Ecopath模型定义生态系统由一系列生态关联的功能组构成, 模型假设每个功能组的能量输入和输出平衡[6, 7]。
模型中的功能组是指生态学或者分类地位上相似的物种的集合, 也可以把单个物种作为独立的功能组[8, 12]。首先对太白河生态系统进行初步划分: 浮游植物、浮游动物、底栖生物及鱼类。然后根据食性特点、生活水层将鱼类进一步划分为川陕哲罗鲑、秦岭细鳞鲑及小型中下层鱼类共3个功能组。各功能组的名称及其包括的主要种类见表 2。
表 2 太白河生态系统功能组的划分Table 2. Division of functional groups in Taibai River ecosystem编号
ID功能组名称
Function group name包含的种类
Category of inclusion1 川陕哲罗鲑
Hucho bleekeri川陕哲罗鲑
Hucho bleekeri2 秦岭细鳞鲑
Brachymystax lenok tsinlingensis秦岭细鳞鲑
Brachymystax lenok tsinlingensis3 小型中下层鱼类
Small lower-top fishes拉氏鱥、汉水高原鳅、中华花鳅
Rhynchocypris lagowskii, Triplophysa shaanxiensis, Siberian spiny loach4 浮游动物
Zooplankton原生动物、桡足类、轮虫、枝角类
Protozoa, Copepods, Rotifers, Cladocera5 浮游植物
Phytoplankton硅藻门、绿藻门等
Bacillariophyta Chlorophyta et al.6 底栖动物
Benthos昆虫纲、腹足纲等
Insecta, Gastropoda et al.7 碎屑
Detritus腐烂的动植物等碎屑
Detritus of decaying animals and plants1.5 生态学参数设置
生物量(B)以湿重表示, 指单位时间内单位面积或单位体积存在的某种生物总量, 在Ecopath模型中的单位为t/km2。
水生植物、底栖动物、鱼类采样量纲转换公式:
1 g/m2=10–6/(10–3 km)2=1 t/km2
浮游动、植物采样量纲转换公式[9]:
1 g/L×M=1 g/dm3×M=10–5 t/(10–4 km)2=103 t/km2
式中, M为水深, 单位为m。
碎屑的生物量由初级生产力(包括水生植物和浮游植物)和水体的透明度之间的关系计算, 计算公式:
lgD=0.954lgPP+0.863lgE–2.41
式中, D为碎屑生物量, g/m2; pp为初级生产量, g/(m2·a), E为平均透明度, m。
生产量与生物量比值(P/B)也称生物量的周转率, 在生态系统平衡状态下, 鱼类功能组的P/B系数等同于自然死亡率, 利用Pauly经验公式进行计算, 具体估计通过FishBase网站上的生活—历史工具得出[13], 其他功能组的P/B系数数值参考中小型河流的相关研究[14—16]。
消费量与生物量比值(Q/B)。鱼类功能组的Q/B值在FishBase网站上参考Palomares和Pauly经验公式估算求出[17], 其他功能组的Q/B系数数值参考中小型河流的相关研究[18]。
各组未同化食物比例(GSi)虽不影响Ecopath模型的营养平衡, 却是估计能量收支必不可少的参数。本研究通过参考相似文献和Ecopath手册设置, 其中肉食性鱼类和杂食性鱼类的未同化食物比例分别设置为0.20和0.41, 浮游动物和底栖动物分别设为0.65和0.91[19]。
食物网矩阵(DCi)用于描述各功能组之间捕食与被捕食的详细数量关系。鱼类功能组的相关食性主要参考Fishbase网站和已有的历史食性研究资料[20—26], 浮游动物与底栖动物的食性参照目前的食性研究结果, 对于包含多种类别的鱼类和底栖动物功能组, 它们的食物组成根据生物量的权重计算得出[27, 28]。
2. 结果
2.1 鱼类资源
共采集到鱼类5种, 隶属于2目3科, 捕获数量最多的为拉氏鱥(Rhynchocypris lagowskii; 51.77%), 其次为汉水高原鳅(Triplophysa shaanxiensis; 28.75%)。川陕哲罗鲑捕获数量最低, 仅占渔获物总数的4.9%。秦岭细鳞鲑和川陕哲罗鲑在栖息地分布上存在明显差异, 秦岭细鳞鲑主要分布在太白河上端, 川陕哲罗鲑则主要分布在中下段, 在上段亦有少量分布(表 3)。
表 3 2017—2018年太白河渔获物组成Table 3. Fish community structure in Taibai River from 2017 to 2018鱼类
Fish拉氏鱥
Rhynchocypris lagowskii汉水高原鳅
Triplophysa shaanxiensis中华花鳅
Siberian spiny loach秦岭细鳞鲑
Brachymystax lenok tsinlingensis川陕哲罗鲑
Hucho bleekeri尾数Mantissa 380 211 49 58 36 占比Account (%) 51.77 28.75 6.68 7.9 4.9 根据密度法, 估算出太白河川陕哲罗鲑的资源总量为3814尾, 95%的置信区间为3624—4005尾。成鱼的资源量为1286尾, 95%的置信区间为1233—1339尾; 幼鱼的资源量为2528尾, 95%的置信区间为2438—2619尾(表 4)。
表 4 太白河川陕哲罗鲑资源量的估算Table 4. Estimates of the resource of the Hucho bleekeri in the Taibai Mountains指标Index 成鱼
Adult fish幼鱼
Juvenile fish总体
Sum样点平均密度(尾/m2)
Sample average density0.0043 0.0084 0.0127 标准差
Standard deviation0.005 0.009 0.019 绝对误差
Absolute error0.0018 0.0030 0.0064 调查精度
Accuracy of survey95.89 96.49 95.01 平均资源量(尾)
Average resources1286 2528 3814 95%上限(尾)
95% upper limit1339 2619 4005 95%下限(尾)
95% lower limit1233 2438 3624 2.2 环境特征与水生生物资源
研究结果显示太白河年平均水温为11.6℃, 范围为0.1—22.5℃; 年平均溶氧为8.76 mg/L, 范围为7.52—9.43 mg/L; 流速为1.69 m/s, 范围为0.88—2.45 m/s。透明度高, 最高可达1.5m以上。
水生生物资源调查结果表明, 浮游动物包括原生动物、桡足类、轮虫类和枝角类。浮游植物以硅藻为主, 占总种数的70.21%。昆虫是底栖动物中数量最多的, 占总种数的82.14% (表 5)。上述饵料生物的生物量与物种组成均较高(表 6)。
表 5 太白河水生生物资源组成Table 5. Composition of aquatic biological resources in the Taibai River种类
Species种类数量
Species number比例
Proportion (%)浮游动物
Zooplankton原生动物Protozoan 7 53.85 桡足类Copepoda 3 23.08 枝角类Rotifer 2 15.38 轮虫Cladocera 1 7.69 浮游植物
Phytoplankton硅藻门Bacillariophyta 33 70.21 绿藻门Chlorophyta 8 17.02 蓝藻门Cyanophyta 2 4.26 金藻门Chrysophyta 2 4.26 黄藻门Xanthophyta 1 2.13 裸藻门Euglenophyta 1 2.13 底栖生物
Benthic animal昆虫纲Insecta 23 82.14 腹足纲Gastropoda 2 7.14 寡毛纲Oligochaeta 1 3.57 涡虫纲Turbellaria 1 3.57 线虫纲Nematoda 1 3.57 表 6 太白河饵料生物生物量Table 6. Seasonal statistics of the biomass of food organisms指标
Index春季
Spring夏季
Summer秋季
Autumn冬季
Winter平均值
Average浮游动物
Zooplankton (mg/L)0.17 0.21 0.16 0.25 0.20 浮游植物
Phytoplankton (mg/L)1.37 2.34 1.62 0.89 1.55 底栖生物
Benthic animal (mg/L)2.88 2.47 2.83 2.83 2.75 2.3 模型估算的输出参数及生态学意义
在模型达到平衡后, 其基本参数如表 7和表 8所示, 加粗数值为模型估算。由于各功能组的生物量积累和迁移量在本研究中均为0, 故生态营养转化效率EE反映的是各功能组生产量被捕食所利用的程度。模型达到平衡后浮游动物、底栖动物、小型中下层鱼类功能组EE值(生态营养转化效率)在0.823—0.911, 利用程度相对较高, 而川陕哲罗鲑、秦岭细鳞鲑、浮游植物和碎屑功能组利用程度不高(EE值分别为0、0、0.223和0.135)。
表 7 太白河Ecopath模型的输入和输出参数Table 7. Input and output parameters of Ecopath model of Taibai River功能组
Functional
group营养级
Trophic
level生物量
Biomass
(t/km2)生产量/
生物量
Production/
Biomass消耗量/
生物量
Consumption/
Biomass生态营养
转化效率
Ecotrophic
efficiency1 3.304 1.391 0.31 5.7 0.000 2 3.256 0.816 0.23 7.0 0.000 3 2.481 3.560 2.60 18.6 0.823 4 2.000 2.000 40 160.0 0.846 5 1.000 15.500 106 0.223 6 2.155 2.750 5.83 109.0 0.911 7 1.000 4.540 0.135 注: 各功能组的种类组成如表 2所示Note: Main species composition of the functional groups is shown in Tab. 2 2.4 太白河生态系统总体特征
太白河生态系统的总生产量为1748.91 t/(km2·a)、总初级生产力为1643.00 t/(km2·a), 约占总生产量的93.94%。该水域生态系统的连接指数(CI)和系统杂食指数(SOI)分别为0.429和0.087。
表 8 太白河生态系统总特征参数Table 8. Total characteristic parameters of Taibai River ecosystem生态系统参数
Parameter数值
Number单位
Unit总消耗量
Sum of all consumption699.61 t/(km2·a) 总输出量
Sum of all exports1559.95 t/(km2·a) 总呼吸量
Sum of all respiratory flows83.05 t/(km2·a) 流向碎屑总量
Sum of all flows into detritus1802.74 t/(km2·a) 系统总流量
Total system throughput4145.34 t/(km2·a) 系统总生产量
Sum of all production1748.91 t/(km2·a) 总净初级生产量
Total net primary production1643.00 t/(km2·a) 总初级生产量/总呼吸量
Total primary production/total respiration(P/R)19.783 系统净生产量
Net system production1599.95 t/km2 总初级生产量/总生物量
Total primary production/total biomass(P/R)63.151 总生物量/总流量
Total biomass/total throughput0.006 系统连接指数
Connectance index(CI)0.429 系统杂食指数
System omnivory index(SOI)0.115 2.5 太白河营养级结构和物质能量流动
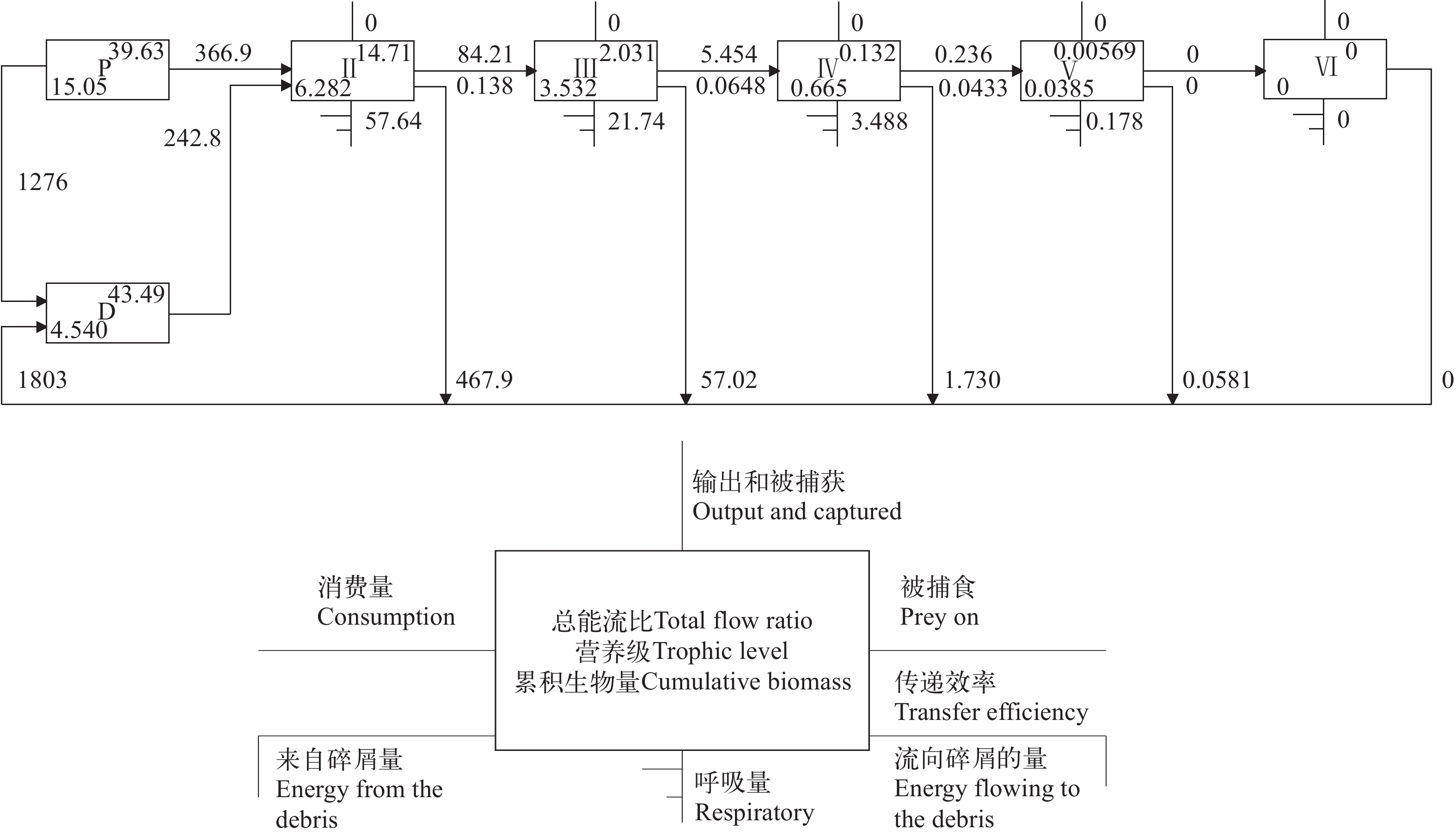
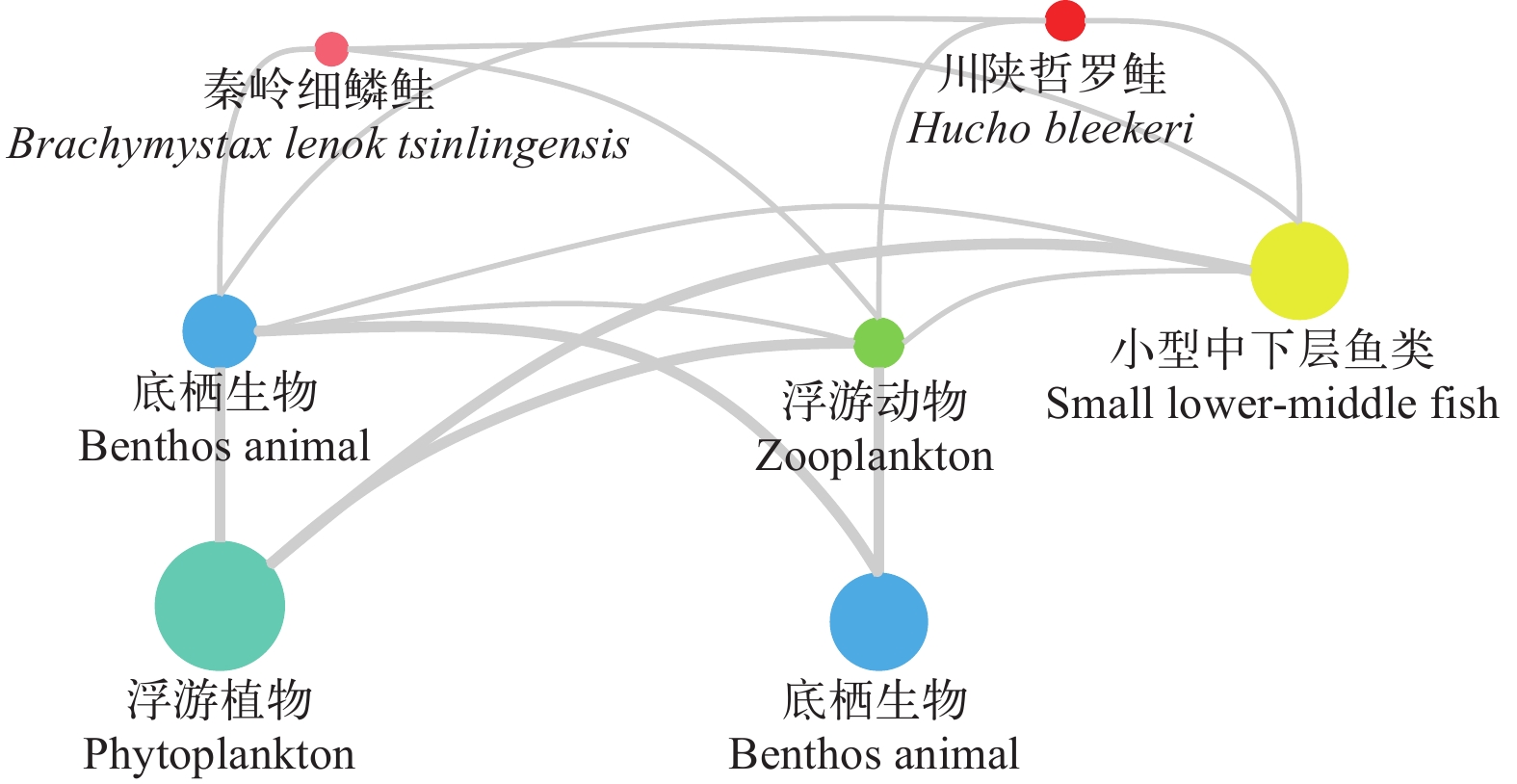
如图 2所示, 秦岭细鳞鲑与川陕哲罗鲑主要占据Ⅲ和Ⅳ营养级, 少量占据Ⅴ营养级。从各离散营养级的具体流量分布来看, 太白河生态系统的营养物质主要在前5个离散营养级间流动。太白河流域食物链的能量流动过程可分为牧食食物链和碎屑食物链(图 3)。在2条食物链合并后, 太白河生态系统营养级Ⅱ、Ⅲ、Ⅳ、Ⅴ的传递效率依次分别为13.8%、6.5%和4.3%呈典型的金字塔分布。
![]() 图 2 太白河生态系统营养级间的物质流动Figure 2. Material flow between trophic levels in the Taibai River ecosystem
图 2 太白河生态系统营养级间的物质流动Figure 2. Material flow between trophic levels in the Taibai River ecosystem2.6 太白河生态系统功能组间营养关系和关键种分析
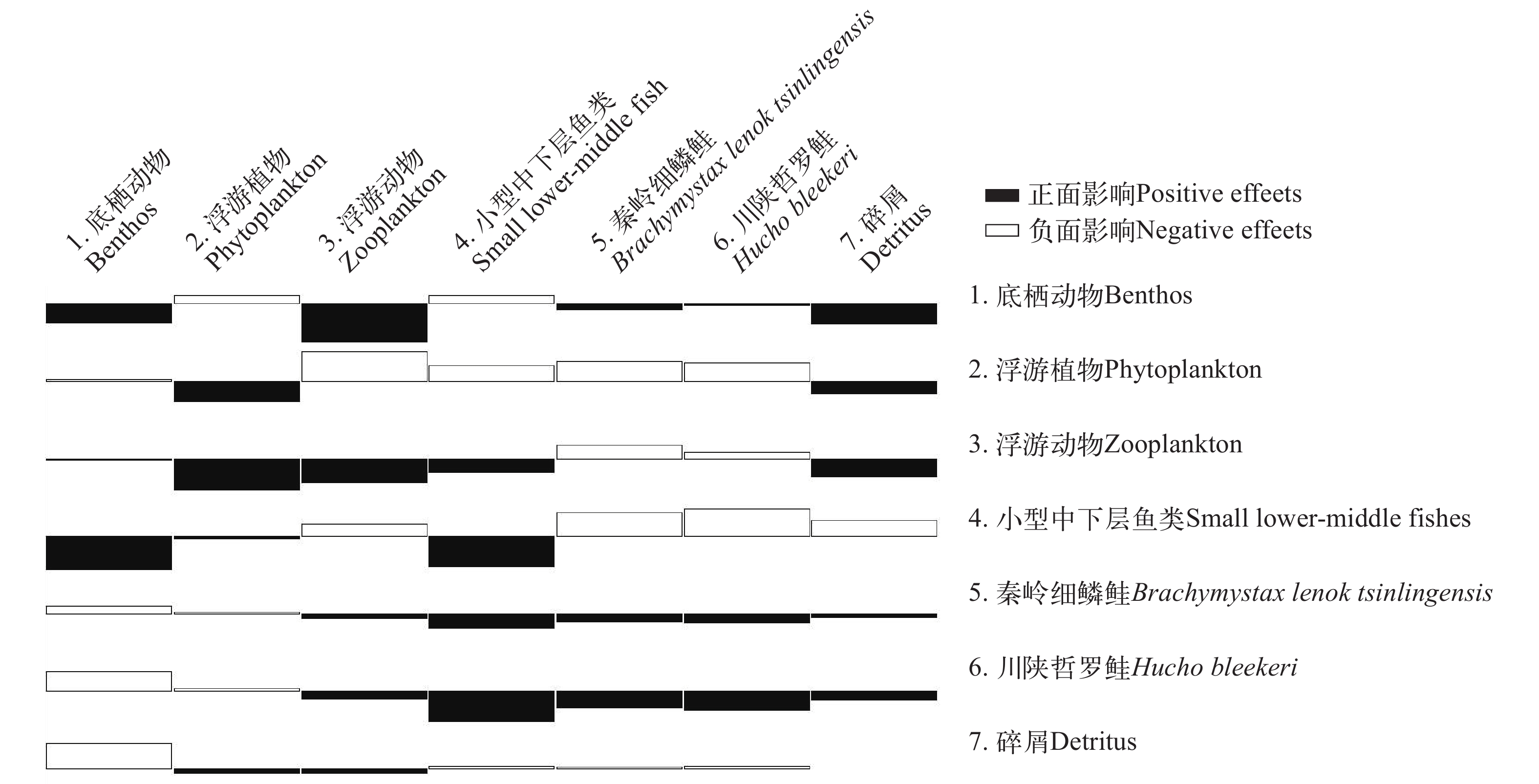
如图 4所示, 浮游植物、浮游动物、小型中下层鱼类对川陕哲罗鲑功能组呈正面影响, 其中浮游植物与小型中下层鱼类影响程度更深。秦岭细鳞鲑与川陕哲罗鲑对川陕哲罗鲑功能组呈负面影响, 其中川陕哲罗鲑功能组自身影响程度更深。太白河生态系统中小型中下层鱼类与川陕哲罗鲑为关键种。
![]() 图 4 太白河生态系统各功能组间的交互营养影响Figure 4. Cross-trophic effects among functional groups in the Taibai River ecosystem
图 4 太白河生态系统各功能组间的交互营养影响Figure 4. Cross-trophic effects among functional groups in the Taibai River ecosystem2.7 太白河川陕哲罗鲑环境容纳量
太白河生态系统Ecopath模型调试和优化至稳定后, 采用逐步提高生态系统中川陕哲罗鲑的种群数量, 以其主要饵料资源功能组的EE作为指标, 当EE大于1时, 模型失去平衡, 此时的种群数量即为该水域川陕哲罗鲑的环境容纳量[29]。
迭代计算发现, 当川陕哲罗鲑种群数量增加到1.87 t/ km2时, 此时小型中下层鱼类功能组EE=1, 同时, 其他功能组的能量流动与生物量没有明显改变, 即为系统的环境容纳量。因此, 推算出太白河生态系统支撑的川陕哲罗鲑平均生物量为1.87 t/km2, 根据渔获调查数据可知为5127尾, 其中幼鱼为3399尾, 成鱼为1728尾。
3. 讨论
3.1 太白河川陕哲罗鲑环境容纳量
受水生生物调查本身局限性的影响, 本研究估算的太白河川陕哲罗鲑环境容纳量可能比真实值低。本次太白河鱼类调查(2017年10月)记录到的鱼类有5种, 其中拉氏鱥数量最多, 占比51.77%, 其次为汉水高原鳅(28.75%), 另有少量川陕哲罗鲑、中华花鳅、秦岭细鳞鲑等物种。同时期对同属汉江上游溪流的湑水河鱼类调查显示, 在2017—2019年开展的6次鱼类调查共记录到鱼类27种[30]。由于太白河没有同期甚至历史的鱼类资源参考数据, 以湑水河的调查结果参考来看, 本次太白河鱼类调查(2017年10月)记录到的鱼类种类应该是偏少的。本研究构建的太白河生态系统中浮游植物EE值相对浮游动物、底栖生物等EE值较低, 为0.223。理论计算显示, 模型中可能缺少草食性鱼类。通过横向对比和理论计算可推测太白河生态系统中鱼类种类和资源量应该高于本次调查结果, 这可能是本次调查采样点以川陕哲罗鲑历史栖息地为核心所导致的结果偏差。考虑到川陕哲罗鲑占据了太白河生态系统中最高营养级, 未调查到的鱼类均可作为川陕哲罗鲑的饵料鱼[31, 32]。而模型显示制约太白河川陕哲罗鲑环境容纳量的关键因素是饵料鱼生物量, 因此可推测太白河川陕哲罗鲑真实的环境容纳量要比本研究的估算值更高。
3.2 太白河水生态系统的稳定性
生态系统总初级生产力(TPP)与总呼吸量(TR)的比值TPP/TR是表征系统成熟度的重要指标, 一般在发育成熟的生态系统中TPP/TR的值接近1, 而发育中的系统该值大于1, 遭受污染的系统中该值小于1[32]。我国北方中小型河流生态系统TPP/TR=3.630[14], 但太白河生态系统的TPP/TR值为19.783, 表明太白河目前仍旧是一个发育中的系统。成熟的系统其功能组间的CI与SOI指数值均接近1[33, 34], 该水域生态系统的CI和SOI指数值为0.429和0.115, 表明该生态系统各功能组之间的聚合度低, 稳定性不高, 相对较脆弱。
此外, 太白河生态系统中浮游动物、底栖生物及小型中下层鱼类功能组的EE值很高, 表明川陕哲罗鲑与秦岭细鳞鲑几乎不存在被捕食与捕捞的压力, 对小型鱼类和幼鱼带来了较大的捕食风险。
本次构建的太白河生态系统主要目的是估算太白河川陕哲罗鲑环境容纳量, 捕捞到的鱼类种类较少, 并未进行系统的鱼类资源调查, 因此生态系统评价结果可能存在一定误差。
3.3 川陕哲罗鲑种群的威胁因素
20世纪60年代以前, 川陕哲罗鲑自然资源量较多, 其分布较广, 几乎遍及岷江上游和大渡河水系, 青衣河、天全河、大川河、玛柯河、太白河都有分布[35]。在60年代以后, 由于人类活动的因素: 拦河筑坝、兴建水利工程、挖沙、开采金矿等, 致使川陕哲罗鲑栖息地的生态环境遭到破坏, 分布区域不断缩小; 过度捕捞、电鱼毒鱼等方面因素导致其自然资源量急剧下降, 这是川陕哲罗鲑种群衰退的主要因素。目前在原有历史栖息地已很难采集到样本[36]。自2012年秋季时隔30年太白河流域再次发现川陕哲罗鲑种群至本次渔获物调查发现川陕哲罗鲑野生幼鱼, 证实了太白河川陕哲罗鲑存在自然繁殖[5]。但通过密度法估算的太白河川陕哲罗鲑资源量为3814尾, 远未达到估算的环境容纳量。除了人为活动, 有学者认为气候变化对鲑科鱼类的自然繁殖有着直接影响, 川陕哲罗鲑作为我国特有的、局部分布的陆封型鲑科鱼类, 气候变化导致的栖息地水温与流速的变化或许是制约其种群恢复的原因之一。有研究表明夏秋季节最高水温的升高会降低幼年鲑的存活率, 而春冬变暖则会提高鲑科鱼类的发育速度[37, 38]。2017—2018年太白河年平均水温为11.6℃, 8月份水温达到最高, 平均19.3℃。川陕哲罗鲑最适生长温度为14.6℃, 2017—2018年全年共有4个月平均水温超过其最适生长温度。鲑科鱼类的产卵时间取决于成年鲑忍受夏季高温的能力以及鱼卵的质量。鲑科鱼类的鱼卵如果长时间暴露于超过8℃的水温中将无法存活[38], 而川陕哲罗鲑的繁殖期为4月中下旬, 根据水温监测数据, 2017—2018年太白河4月底至五月初平均水温超过8℃。因此气候变化导致水温上升可能是影响川陕哲罗鲑自然繁殖的原因之一, 但仍需要针对性研究来证实。
气候变化导致的我国汛期提前且极端气候事件频发也可能是影响鲑科鱼类自然繁殖的主要原因之一。有证据表明, 在20世纪90年代, 额外的湿润泉水在北爱尔兰造成了更高的河流流量, 导致大西洋鲑鱼存活率降低[39]。川陕哲罗鲑产卵方式为筑巢产卵, 近年来太白河5月初进入主汛期与川陕哲罗鲑产卵期接近, 汛期导致的太白河流量增大可能导致川陕哲罗鲑卵巢被冲毁影响其自然繁殖。综上所述, 气候变化对鲑科鱼类的自然繁殖存在一定的影响, 但是否是阻碍川陕哲罗鲑自然种群恢复的主要原因之一仍需要针对性的研究与实验来证实。
3.4 川陕哲罗鲑的保护建议
良好的栖息环境对野生生物资源的恢复是至关重要的。本研究调查发现, 太白河流域存在陕西太白黄金矿业, 存在明显水域环境污染源。此外, 太白河流域修建了2个水电站, 存在水电开发和水坝修建等工程, 河道内泥沙淤积。这些人为活动导致川陕哲罗鲑适宜的栖息环境被破坏, 不利于川陕哲罗鲑的种群恢复。因此需要加强保护陆地生境, 提高植被蓄水能力, 降低瞬时洪峰, 从而提高川陕哲罗鲑产卵繁殖的容量。同时应在全面禁捕的大背景下落实太白河环境保护及禁捕工作, 降低人为活动与气候变化对川陕哲罗鲑野生种群自然繁殖的影响。
不少学者对川陕哲罗鲑适宜的生境和产卵条件进行研究, 处于不同生活史时期的川陕哲罗鲑对河流生境需求不同。川陕哲罗鲑栖息地环境特点为水深较深, 河流透明度高, 水流湍急, 河谷呈现V型。王凤等[40]分析和比较几种鲑科鱼类的繁殖特性, 其中川陕哲罗鲑的适宜水温为4—14℃, 表明温度对川陕哲罗鲑分布具有重大影响。本研究调查发现, 太白河流域森林覆盖率高, 水温在1月时最低, 水温为2.0℃, 随后逐月升高, 在8月达到最高, 水温为19.3℃, 年平均水温为11.6℃, 太白河平均溶氧为8.76 mg/L, 溶氧含量丰富, 水体呈弱碱性, 水体流速较快, 水体清澈透明, 使得川陕哲罗鲑自然种群得以保留。栖息地破坏会将区域内物种分隔为局部物种, 最终形成小种群, 若局部种群栖息地被破坏, 最终导致局部种群灭亡[41, 42]。自20世纪以来, 人为活动导致川陕哲罗鲑栖息地分布范围缩减, 资源量呈下降趋势。这些栖息地的生态环境已经发生了巨大的变迁, 可能已不再适合川陕哲罗鲑的生长繁殖。因此, 建议开展太白河川陕哲罗鲑产卵场与繁殖条件的调查, 针对现存产卵场制定保护措施, 以促进川陕哲罗鲑自然种群资源的恢复。
4. 结论
本研究表明, 太白河川陕哲罗鲑野生种群现存量为3814尾, 其中成鱼的资源量为1286尾, 幼鱼的资源量为2528尾。初步估算的太白河川陕哲罗鲑环境容纳量为5127尾, 实际环境容纳量应该高于估算量, 建议通过保护陆地生境, 提高植被蓄水能力, 降低瞬时洪峰, 提高川陕哲罗鲑产卵繁殖的容量以及增殖放流的手段恢复川陕哲罗鲑野生种群, 同时加强栖息地保护, 促进川陕哲罗鲑资源恢复。
-
![]()
图 2 太白河生态系统营养级间的物质流动
Figure 2. Material flow between trophic levels in the Taibai River ecosystem
![]()
图 4 太白河生态系统各功能组间的交互营养影响
Figure 4. Cross-trophic effects among functional groups in the Taibai River ecosystem
表 1 太白河基本环境特征调查内容
Table 1 Basic environmental characteristics of the Taibai River survey content
类别
Category调查内容
Investigation content断面及频次
Section and frequency数据来源
Data source调查方法
Investigation method流域特性
Characteristics of river basin流量、流域面积、比降、植被覆盖、人口 整体1次 文献 通过查阅地方志、统计年鉴等资料获得 水体理化特性
Physicochemical characteristics水温、溶氧、流速、透明度、pH 在A段和C段分别设置1个和2个断面, 按照季节调查4次 实地测量 采用美国YSI公司 Proplus 便携式水质监测仪进行测量 水生生物现状
Aquatic life status鱼类、浮游动植物、底栖生物、水生高等植物 与水体理化特性相同 实地采集 参考《河流水生态调查技术方法》[11]  下载: 导出CSV
下载: 导出CSV
表 2 太白河生态系统功能组的划分
Table 2 Division of functional groups in Taibai River ecosystem
编号
ID功能组名称
Function group name包含的种类
Category of inclusion1 川陕哲罗鲑
Hucho bleekeri川陕哲罗鲑
Hucho bleekeri2 秦岭细鳞鲑
Brachymystax lenok tsinlingensis秦岭细鳞鲑
Brachymystax lenok tsinlingensis3 小型中下层鱼类
Small lower-top fishes拉氏鱥、汉水高原鳅、中华花鳅
Rhynchocypris lagowskii, Triplophysa shaanxiensis, Siberian spiny loach4 浮游动物
Zooplankton原生动物、桡足类、轮虫、枝角类
Protozoa, Copepods, Rotifers, Cladocera5 浮游植物
Phytoplankton硅藻门、绿藻门等
Bacillariophyta Chlorophyta et al.6 底栖动物
Benthos昆虫纲、腹足纲等
Insecta, Gastropoda et al.7 碎屑
Detritus腐烂的动植物等碎屑
Detritus of decaying animals and plants
下载: 导出CSV
表 3 2017—2018年太白河渔获物组成
Table 3 Fish community structure in Taibai River from 2017 to 2018
鱼类
Fish拉氏鱥
Rhynchocypris lagowskii汉水高原鳅
Triplophysa shaanxiensis中华花鳅
Siberian spiny loach秦岭细鳞鲑
Brachymystax lenok tsinlingensis川陕哲罗鲑
Hucho bleekeri尾数Mantissa 380 211 49 58 36 占比Account (%) 51.77 28.75 6.68 7.9 4.9
下载: 导出CSV
表 4 太白河川陕哲罗鲑资源量的估算
Table 4 Estimates of the resource of the Hucho bleekeri in the Taibai Mountains
指标Index 成鱼
Adult fish幼鱼
Juvenile fish总体
Sum样点平均密度(尾/m2)
Sample average density0.0043 0.0084 0.0127 标准差
Standard deviation0.005 0.009 0.019 绝对误差
Absolute error0.0018 0.0030 0.0064 调查精度
Accuracy of survey95.89 96.49 95.01 平均资源量(尾)
Average resources1286 2528 3814 95%上限(尾)
95% upper limit1339 2619 4005 95%下限(尾)
95% lower limit1233 2438 3624
下载: 导出CSV
表 5 太白河水生生物资源组成
Table 5 Composition of aquatic biological resources in the Taibai River
种类
Species种类数量
Species number比例
Proportion (%)浮游动物
Zooplankton原生动物Protozoan 7 53.85 桡足类Copepoda 3 23.08 枝角类Rotifer 2 15.38 轮虫Cladocera 1 7.69 浮游植物
Phytoplankton硅藻门Bacillariophyta 33 70.21 绿藻门Chlorophyta 8 17.02 蓝藻门Cyanophyta 2 4.26 金藻门Chrysophyta 2 4.26 黄藻门Xanthophyta 1 2.13 裸藻门Euglenophyta 1 2.13 底栖生物
Benthic animal昆虫纲Insecta 23 82.14 腹足纲Gastropoda 2 7.14 寡毛纲Oligochaeta 1 3.57 涡虫纲Turbellaria 1 3.57 线虫纲Nematoda 1 3.57
下载: 导出CSV
表 6 太白河饵料生物生物量
Table 6 Seasonal statistics of the biomass of food organisms
指标
Index春季
Spring夏季
Summer秋季
Autumn冬季
Winter平均值
Average浮游动物
Zooplankton (mg/L)0.17 0.21 0.16 0.25 0.20 浮游植物
Phytoplankton (mg/L)1.37 2.34 1.62 0.89 1.55 底栖生物
Benthic animal (mg/L)2.88 2.47 2.83 2.83 2.75
下载: 导出CSV
表 7 太白河Ecopath模型的输入和输出参数
Table 7 Input and output parameters of Ecopath model of Taibai River
功能组
Functional
group营养级
Trophic
level生物量
Biomass
(t/km2)生产量/
生物量
Production/
Biomass消耗量/
生物量
Consumption/
Biomass生态营养
转化效率
Ecotrophic
efficiency1 3.304 1.391 0.31 5.7 0.000 2 3.256 0.816 0.23 7.0 0.000 3 2.481 3.560 2.60 18.6 0.823 4 2.000 2.000 40 160.0 0.846 5 1.000 15.500 106 0.223 6 2.155 2.750 5.83 109.0 0.911 7 1.000 4.540 0.135 注: 各功能组的种类组成如表 2所示Note: Main species composition of the functional groups is shown in Tab. 2
下载: 导出CSV
表 8 太白河生态系统总特征参数
Table 8 Total characteristic parameters of Taibai River ecosystem
生态系统参数
Parameter数值
Number单位
Unit总消耗量
Sum of all consumption699.61 t/(km2·a) 总输出量
Sum of all exports1559.95 t/(km2·a) 总呼吸量
Sum of all respiratory flows83.05 t/(km2·a) 流向碎屑总量
Sum of all flows into detritus1802.74 t/(km2·a) 系统总流量
Total system throughput4145.34 t/(km2·a) 系统总生产量
Sum of all production1748.91 t/(km2·a) 总净初级生产量
Total net primary production1643.00 t/(km2·a) 总初级生产量/总呼吸量
Total primary production/total respiration(P/R)19.783 系统净生产量
Net system production1599.95 t/km2 总初级生产量/总生物量
Total primary production/total biomass(P/R)63.151 总生物量/总流量
Total biomass/total throughput0.006 系统连接指数
Connectance index(CI)0.429 系统杂食指数
System omnivory index(SOI)0.115
下载: 导出CSV
-
[1] 邓小林. 玛可河水域川陕哲罗鲑的濒危原因与保护措施 [J]. 甘肃农业, 2007(6): 87-88. doi: 10.3969/j.issn.1673-9019.2007.06.039 Deng X L. Endangerment reasons and protection measures of Hucho bleekeri in the Maco River [J]. Gansu Agriculture, 2007(6): 87-88. doi: 10.3969/j.issn.1673-9019.2007.06.039
[2] 王成友, 杜浩, 吴金明, 等. 光照对川陕哲罗鲑受精卵孵化的影响 [J]. 淡水渔业, 2019, 49(1): 62-65. doi: 10.3969/j.issn.1000-6907.2019.01.011 Wang C Y, Du H, Wu J M, et al. Effect of illumination on the hatching of Sichuan taimen (Hucho bleekeri Kimura) eggs [J]. Freshwater Fisheries, 2019, 49(1): 62-65. doi: 10.3969/j.issn.1000-6907.2019.01.011
[3] Zhang S H, Wei Q W, Du H, et al. The complete mitochondrial genome of the endangered Hucho bleekeri (Salmonidae: Huchen) [J]. Mitochondrial DNA Part A,DNA Mapping,Sequencing,and Analysis, 2016, 27(1): 124-125.
[4] 丁瑞华, 卿足平. 虎嘉鱼保护生物学的研究Ⅰ. 分布区域及其变迁 [J]. 四川动物, 1994, 13(4): 152-154. Ding R H, Qing Z P. Protection biology of Hucho bleekeri Ⅰ. Distribution region and its changes [J]. Sichuan Journal of Zoology, 1994, 13(4): 152-154.
[5] 杜浩, 李罗新, 危起伟, 等. 濒危物种川陕哲罗鲑在汉江上游太白河再发现 [J]. 动物学杂志, 2014, 49(3): 414. Du H, Li L X, Wei Q W, et al. The rediscovery of Hucho bleekeri in the Taibai River, the upper tributary of the Hanjiang River, China [J]. Chinese Journal of Zoology, 2014, 49(3): 414.
[6] Christensen V, Walters C, Pauly D, et al. Ecopath with Ecosim version 6 [J]. 2008.
[7] 李永涛. 天鹅洲江豚生境选择, 环境容纳量和种群生存力分析——迁地保护理论初探 [D]. 北京: 中国科学院大学, 2017: 33-39. Li Y T. Analysis of habitat selection, environmental capacity and population viability of Tianezhou finless porposie preliminary study on ex situ conservation theory [D]. Beijing: University of Chinese Academy of Sciences, 2017: 33-39.
[8] 徐超, 王思凯, 赵峰, 等. 基于Ecopath模型的长江口生态系统营养结构和能量流动研究 [J]. 海洋渔业, 2018, 40(3): 309-318. doi: 10.3969/j.issn.1004-2490.2018.03.006 Xu C, Wang S K, Zhao F, et al. Trophic structure and energy flow of the Yangtze Estuary ecosystem based on the analysis with Ecopath model [J]. Marine Fisheries, 2018, 40(3): 309-318. doi: 10.3969/j.issn.1004-2490.2018.03.006
[9] 杨林林, 姜亚洲, 袁兴伟, 等. 象山港典型增殖种类的生态容量评估 [J]. 海洋渔业, 2016, 38(3): 273-282. doi: 10.3969/j.issn.1004-2490.2016.03.006 Yang L L, Jiang Y Z, Yuan X W, et al. Ecological carrying capacity of typical enhancement species in Xiangshan Bay [J]. Marine Fisheries, 2016, 38(3): 273-282. doi: 10.3969/j.issn.1004-2490.2016.03.006
[10] 吴斌, 贺刚, 王伟萍. 基于Ecopath 模型的鄱阳湖通长江水道长江江豚环境容纳量初步估算 [J]. 江西农业学报, 2022, 34(5): 155-159. Wu B, He G, Wang W P. Preliminary estimation of environmental capacity of Yangtze finless porpoise in Poyang Lake through Yangtze River waterway based on ecopath model [J]. Acta Agriculturae Jiangxi, 2022, 34(5): 155-159.
[11] 孟伟, 张远, 渠晓东, 等. 河流生态调查技术方法 [M]. 北京: 科学出版社, 2011: 92-123. Meng W, Zhang Y, Qu X D, et al. Techniques and Methods of River Ecological Survey [M]. Beijing: Science Press, 2011: 92-123.
[12] 张远, 赵长森, 杨胜天, 等. 基于Ecopath的小清河河流生态系统关键功能组分析 [J]. 南水北调与水利科技, 2017, 15(6): 66-73. Zhang Y, Zhao C S, Yang S T, et al. Analysis of key functional group for Xiaoqinghe River ecosystem based on Ecopath [J]. South-to-North Water Transfers and Water Science & Technology, 2017, 15(6): 66-73.
[13] Lima M A L, Doria C R, Carvalho A R, et al. Fisheries and trophic structure of a large tropical river under impoundment [J]. Ecological Indicators, 2020(113): 106-162.
[14] 鲁浩天, 郝子垚, 刘萌硕, 等. 基于Ecopath模型的北方典型中小河流生态系统营养结构与能量流动分析 [J]. 生态与农村环境学报, 2023, 39(11): 1453-1463. Lu H T, Hao Z Y, Liu M S, et al. Analysis on the ecosystem trophic structure and energy flow of typical small and medium rivers in northern China on Ecopath model [J]. Journal of Ecology and Rural Environment, 2023, 39(11): 1453-1463.
[15] 李云凯, 刘恩生, 王辉, 等. 基于Ecopath模型的太湖生态系统结构与功能分析 [J]. 应用生态学报, 2014, 25(7): 2033-2040. Li Y K, Liu E S, Wang H, et al. Analysis on the structure and function of Lake Taihu based on Ecopath model [J]. Chinese Journal of Applied Ecology, 2014, 25(7): 2033-2040.
[16] 张朋飞, 崔晓宇, 骆辉煌, 等. 莲石湖生态系统营养结构与能量流动分析 [J]. 海洋湖沼通报, 2021, 43(6): 124-132. Zhang P F, Cui X Y, Luo H H, et al. Trophic structure and energy flow of Lianshi Lake ecosystem [J]. Transactions of Oceanology and Limnology, 2021, 43(6): 124-132.
[17] Palomares M L D, Pauly D. Predicting food consumption of fish populations as functions of mortality, food type, morphometrics, temperature and salinity [J]. Marine and Freshwater Research, 1998, 49(5): 447. doi: 10.1071/MF98015
[18] 唐剑锋, 肖新宗, 王英才, 等. 南水北调中线干渠生态系统结构与功能分析 [J]. 中国环境科学, 2020, 40(12): 5391-5402. doi: 10.3969/j.issn.1000-6923.2020.12.035 Tang J F, Xiao X Z, Wang Y C, et al. Ecosystem structure and function of the main channel of the middle route of south-north water diversion project [J]. China Environmental Science, 2020, 40(12): 5391-5402. doi: 10.3969/j.issn.1000-6923.2020.12.035
[19] 贾佩峤, 胡忠军, 武震, 等. 基于ecopath模型对滆湖生态系统结构与功能的定量分析 [J]. 长江流域资源与环境, 2013, 22(2): 189-197. Jia P Q, Hu Z J, Wu Z, et al. Quantitative analysis on the structure and function of the Gehu Lake ecosystem based on Ecopath modeling [J]. Resources and Environment in the Yangtze Basin, 2013, 22(2): 189-197.
[20] 武震, 贾佩峤, 胡忠军, 等. 基于Ecopath模型分析分水江水库生态系统结构和功能 [J]. 应用生态学报, 2012, 23(3): 812-818. Wu Z, Jia P J, Hu Z J, et al. Structure and function of Fenshuijiang Reservoir ecosystem based on the analysis with Ecopath model [J]. Chinese Journal of Applied Ecology, 2012, 23(3): 812-818.
[21] 马凯, 佟广香, 张澜澜, 等. 哲罗鲑和川陕哲罗鲑群体多变量形态特征比较[J]. 中国水产科学, 2023, 30(1): 1-10. Ma K, Tong G X, Zhang L L, et al. Multivariate analysis of Hucho taimen and Hucho bleekeri populations based on morphological characteristics [J]. Journal to Fishery Sciences of China, 2023, 30(1): 1-10.
[22] 张建禄, 许涛清, 黄吉芹, 等. 秦岭细鳞鲑生物学参数研究及其个体变化探讨 [J]. 水生态学杂志, 2022, 43(5): 117-121. Zhang J L, Xu T Q, Huang J Q, et al. Biology and miniaturization of Brachymystax lenok tsinlingensis [J]. Journal of Hydroecology, 2022, 43(5): 117-121.
[23] 苟妮娜, 靳铁治, 张建禄, 等. 黑河国家级自然保护区秦岭细鳞鲑主要饵料生物——拉氏鱥种群特征及其季节变化 [J]. 西北农业学报, 2018, 27(9): 1258-1264. doi: 10.7606/j.issn.1004-1389.2018.09.004 Gou N N, Jin T Z, Zhang J L, et al. Population characteristics and seasonal variation of Brachymystax lenoktsinlingensis’s main bait organism Lagowskiella lagowskii in National Nature Reserves of rare aquatic wildlife of Heihe River [J]. Acta Agriculturae Boreali-occidentalis Sinica, 2018, 27(9): 1258-1264. doi: 10.7606/j.issn.1004-1389.2018.09.004
[24] 许鹏辉, 邢顺林, 王小勇, 等. 西藏短尾高原鳅食性组成及rRNA基因ITS间隔分析 [J]. 高原科学研究, 2019, 3(3): 9-14. Xu P H, Xing S L, Wang X Y, et al. Analysis on the diet composition and rRNA gene ITS interval of Trilophysa brevviuda in Tibet [J]. Plateau Science Research, 2019, 3(3): 9-14.
[25] 邓华堂, 岳兴建, 陈大庆, 等. 怒江细尾高原鳅生长特征与食性 [J]. 淡水渔业, 2010, 40(1): 26-33. doi: 10.3969/j.issn.1000-6907.2010.01.005 Deng H T, Yue X J, Chen D Q, et al. Growth characteristics and feed habit of Triplophysa stenura in Nujiang River [J]. Freshwater Fisheries, 2010, 40(1): 26-33. doi: 10.3969/j.issn.1000-6907.2010.01.005
[26] 杨骏, 何兴恒, 孙治宇. 中华花鳅的生长和繁殖生物学的研究 [J]. 水产科学, 2020, 39(2): 209-217. Yang J, He X H, Sun Z Y. Growth and reproductive biology of Chinese spined loach Cobitis sinensis [J]. Fisheries Science, 2020, 39(2): 209-217.
[27] 李昌, 张新, 赵龙, 等. 基于Ecopath模型的密云水库生态系统结构与物质流动特征 [J]. 生物资源, 2021, 43(3): 292-302. Li C, Zhang X, Zhao L, et al. Ecosystem structure and material flows of Miyun Reservoir based on the Ecopath model [J]. Biotic Resources, 2021, 43(3): 292-302.
[28] 陈薛伟杰. 三峡库区万州江段和小江鱼类食物网结构与功能研究 [D]. 贵阳: 贵州大学, 2018: 28-32. Chen X W J. Studies on fish food web structure and function of Wanzhou Section and Xiaojiang River in the Three Gorges Reservoir [D]. Guiyang: Guizhou University, 2018: 28-32
[29] Darwall W R T, Allison E H, Turner G F, et al. Lake of flies, or lake of fish? A trophic model of Lake Malawi [J]. Ecological Modelling, 2010, 221(4): 713-727. doi: 10.1016/j.ecolmodel.2009.11.001
[30] 王慧, 张建新, 田翊霏, 等. 湑水河国家级水产种质资源保护区鱼类资源调查与评价 [J]. 家畜生态学报, 2021, 42(11): 54-59. doi: 10.3969/j.issn.1673-1182.2021.11.011 Wang H, Zhang J X, Tian Y F, et al. Investigation and evaluation of fish resources in the national aquatic germplasm resources conservation zone of Xushui River [J]. Journal of Domestic Animal Ecology, 2021, 42(11): 54-59. doi: 10.3969/j.issn.1673-1182.2021.11.011
[31] 丁瑞华, 桂林华, 李明, 等. 虎嘉鱼及其保护生物学 [J]. 广西师范大学学报(自然科学版), 2010, 28(2): 96-102. Ding R H, Gui L H, Li M, et al. Protective biology of Hucho bleekeri in the upper Changjiang River, China [J]. Journal of Guangxi Normal University (Natural Science Edition), 2010, 28(2): 96-102.
[32] 吴万荣. 大川河水生生物与虎嘉鱼的生态位 [J]. 鲑鳟渔业, 1989, 2(1): 23-33. Wu W R. Ecological niche of Hucho bleekeri kimura and aquatic lifes in Dachuan River [J]. Chinese Journal of Fisheries, 1989, 2(1): 23-33.
[33] Hd J, Odum E P. Fundamentals of ecology [J]. Population (French Edition), 1974, 29(2): 376. doi: 10.2307/1530838
[34] Christensen V, Pauly D. Fish production, catches and the carrying capacity of the world oceans [J]. Naga, 1995, 18(3): 34-40.
[35] 申志新, 唐文家, 李柯懋. 川陕哲罗鲑的生存危机与保护对策 [J]. 淡水渔业, 2005, 35(4): 25-28. doi: 10.3969/j.issn.1000-6907.2005.04.007 Shen Z X, Tang W J, Li K M. Survival crisis and protection countermeasures of Hcho bleekeri in Sichuan and Shaanxi [J]. Freshwater Fisheries, 2005, 35(4): 25-28. doi: 10.3969/j.issn.1000-6907.2005.04.007
[36] 祁得林, 晁燕, 杨成, 等. 川陕哲罗鲑Cyt b基因克隆及其在鲑亚科中的系统发育关系 [J]. 四川动物, 2009, 28(6): 805-809. doi: 10.3969/j.issn.1000-7083.2009.06.002 Qi D L, Chao Y, Yang C, et al. Cloning of mitochondrial cytochrome b gene of Hucho bleekeri and its phylogenetic relationships in subfamily salmoninae [J]. Sichuan Journal of Zoology, 2009, 28(6): 805-809. doi: 10.3969/j.issn.1000-7083.2009.06.002
[37] 梁雄伟, 于洪贤, 刘伟. 气候变化对鲑鳟鱼类栖息环境及相关生物学的影响 [J]. 水产学杂志, 2011, 24(2): 63-68. doi: 10.3969/j.issn.1005-3832.2011.02.014 Liang X W, Yu H X, Liu W. Impacts of climate changes on habitat environment and correlative biology of salmons and trouts [J]. Chinese Journal of Fisheries, 2011, 24(2): 63-68. doi: 10.3969/j.issn.1005-3832.2011.02.014
[38] 王志萍, 李家薰, Fillion D, 等. 气候变化对鲑鱼丰富度和繁殖影响的研究进展 [J]. 海洋科学, 2019, 43(4): 98-104. doi: 10.11759/hykx20180929001 Wang Z P, Li J X, Fillion D, et al. Research progress on the effects of climate change on salmon abundance and reproduction [J]. Marine Sciences, 2019, 43(4): 98-104. doi: 10.11759/hykx20180929001
[39] Walsh C, Kilsby C G. Implications of climate change on flow regime affecting Atlantic salmon [J]. Final Report, 1998: 28-32.
[40] 王凤, 张永泉, 尹家胜. 川陕哲罗鱼、太门哲罗鱼及石川哲罗鱼的生物学比较 [J]. 水产学杂志, 2009, 22(1): 59-63. doi: 10.3969/j.issn.1005-3832.2009.01.014 Wang F, Zhang Y Q, Yin J S. A preliminary comparison of the biology character among Hucho breeker, Hucho taimen and Hucho ishikawai Mori [J]. Chinese Journal of Fisheries, 2009, 22(1): 59-63. doi: 10.3969/j.issn.1005-3832.2009.01.014
[41] 杨德国, 危起伟, 李绪兴, 等. 秦岭湑水河太白段珍稀水生动物分布现状及保护对策 [J]. 中国水产科学, 1999, 6(3): 124-126. Yang D G, Wei Q W, Li X X, et al. The distributing actuality and protecting countermeasure of rare aquatic animals in Xushui River of Qinling Mountains [J]. Journal of Fishery Sciences of China, 1999, 6(3): 124-126.
[42] 乐佩琦, 陈宜瑜, 中国濒危动物红皮书: 鱼类 [M]. 北京: 科学出版社, 1998. Le P Q, Chen Y Y, Red Book of Endangered Animals of China: Fishes [M]. Beijing: Science Press, 1998.
计量
- 文章访问数: 204
- HTML全文浏览量: 39
- PDF下载量: 68