
COMPARATIVE ANALYSIS OF THE GROWTH AND FOOD COMPOSITION OF ODONTOBUTIS OBSCURA IN RICE-FISH AND POND CULTURE MODES
-
摘要:
为探究沙塘鳢(Odontobutis obscura)多元化的养殖模式, 开展了稻田养殖和池塘养殖的对比试验, 比较分析养殖模式对沙塘鳢的生长指标及食物组成的影响, 并通过构建沙塘鳢和饵料生物的相关性方程, 探究了沙塘鳢与饵料生物之间的关系。结果显示, 养殖模式对沙塘鳢的存活率没有显著影响, 但稻田养殖沙塘鳢的存活率略高于池塘养殖沙塘鳢, 且稻田养殖沙塘鳢的体增重(BWG)、相对增重率(RWG)及湿重特定生长率(SGRW)均高于池塘养殖沙塘鳢; 虽然稻田养殖沙塘鳢的摄食率和胃充塞度均低于池塘养殖沙塘鳢, 但其生长指标优于池塘养殖沙塘鳢, 且除青虾(Macrobrachium nipponense)外, 3次采样中稻田养殖沙塘鳢消化道中的饵料生物种类和出现率均多于池塘养殖沙塘鳢。综上, 相较于池塘养殖沙塘鳢, 稻田养殖沙塘鳢的生长表现更佳, 且稻田可以为沙塘鳢提供更丰富的食物资源, 有效降低养殖的成本。沙塘鳢体长与体重呈幂函数关系, 其方程为W=3.0×10–5L2.9183(R2=0.879; n=146; P<0.05), 属于异速生长; 沙塘鳢的口裂宽与其体长及青虾额剑长呈线性相关, 青虾体长与其额剑长、沙塘鳢体长也呈线性相关。
Abstract:In order to investigate the effect of different culture modes on the growth index and food composition of Odontobutis obscura, and to explore the relationship between O. obscura and bait organisms, we compared the rice-fish culture and pond culture of O. obscura, and the results showed that there was no significant effect of culture mode on the survival rate of O. obscura, which in rice-fish culture was slightly higher than that in pond culture. The body weight gain (BWG), relative weight gain rate (RWG) and wet weight specific growth rate (SGRW) of O. obscura in rice-fish culture were all higher than that of pond-cultured O. obscura. The feeding rate and gastric congestion of rice-fish cultured O. obscura were lower than those of pond-cultured O. obscura, but their growth indicators were better. In addition, except for Macrobrachium nipponense, there were more species and occurrence of bait organisms in the digestive tract of rice-fish cultured O. obscura than pond-cultured O. obscura in all three samples. In summary, compared with pond culture of O. obscura, the growth performance of rice-fish culture of O. obscura is better, and the rice-fish culture system can provide more abundant food resources for O. obscura and effectively reduce the cost of breeding. The body length of O.obscura was related to the body weight in a power function with the equation W=3.0×10–5 L2.9183 (R²=0.879; n=146; P<0.05), which was allometric growth. The gape width of O. obscura was linearly related to its body length and the frontal sword length of M. nipponense, and the body length of M. nipponense was also linearly correlated with its frontal sword length and the body length of O. obscura.
-
Keywords:
- Culture modes /
- Growth indicators /
- Food composition /
- Correlation equation /
- Odontobutis obscura
-
稻田养殖和池塘养殖是我国淡水养殖业典型的养殖模式[1]。稻田养殖模式是一种将养殖业与种植业相结合, 达到提高稻田利用率, 增加养殖经济效益的生态农业养殖模式[2]。一方面, 稻田各组分可通过生态协同作用充分满足鱼类捕食和生长的需求而不需要添加其他饵料[3], 节约了养殖的经济成本; 另一方面, 稻田养殖过程中减少了对农药和化肥的使用, 对于改善农田生态环境也具有积极作用[4]。池塘养殖模式是指利用池塘进行水生动植物养殖的生产方式, 具有人工可控度高、水体利用率高等特点[1]。在池塘养殖过程中, 仅仅依靠水体中原有资源很难满足养殖生物的生长需求, 通常需要进行人为干预, 如肥水或投喂配合饲料来满足鱼类生长需要, 但也因此增加了养殖水体污染的风险。养殖模式影响水产动物生长和品质, 其根本原因在于养殖模式对水产动物的生存环境有着直接影响。而生存环境是影响水产动物摄食和生长的关键因素之一。环境中所存在的猎物的种类和数量会限制养殖生物的捕食, 继而影响其生长[5]。养殖生物自身的形态特征如口裂、体长等也会对捕食产生影响[6]。
沙塘鳢(Odontobutis obscura)属鲈形目(Perciformes)、塘鳢科(Eleotridae), 又称沙乌鳢、虎头鱼等, 是一类小型底栖肉食性鱼类, 具有含肉量高、刺少、肉质鲜嫩、氨基酸含量高于多种经济鱼类(如草鱼Ctenopharyngodon idella、鲢Hypophthalmichthys molitrix和鳙Aristichthys nobili等)等特点, 是江浙沪一带的传统名贵鱼类[7, 8]。沙塘鳢主要生活在河沟及湖泊中具有较多水草或泥沙的底层, 游泳能力较弱。育苗阶段喜欢摄食轮虫、枝角类及底栖水生昆虫幼体等, 成鱼阶段则以水中体型较小的鱼虾和软体动物为食[9]。由于青虾(Macrobrachium nipponense)生长快, 经济价值高等优点[10], 本试验将适量的抱卵青虾放入养殖稻田和养殖池塘中, 在为沙塘鳢提供食物的同时, 也可以提升青虾的品质。目前, 国内关于沙塘鳢的研究报道并不多, 针对沙塘鳢的养殖模式也比较单一。本研究通过对比稻田养殖和池塘养殖两种模式对沙塘鳢生长与食物组成的影响, 旨在比较沙塘鳢两种养殖模式的差异; 通过构建相关性方程, 探究沙塘鳢与其饵料生物之间的关系, 为沙塘鳢捕食方面的研究提供更多的理论依据。
1. 材料与方法
1.1 试验地点及试验鱼
本试验于2022年8月9日到12月7日在江苏省苏州市太仓市浏河镇名优水产养殖基地开展。共设有6个试验点, 其中3个为养殖稻田, 其余3个为养殖池塘。每个养殖的稻田面积为10亩, 稻田四周具有环沟, 宽度为3.5 m, 水深1 m; 稻田内部有3条纵沟, 长40 m, 宽3 m, 水深0.8 m。试验期间稻田水位保持为0.2 m, 从而保证沙塘鳢可活动范围为环沟、纵沟及稻田。试验稻田中水稻品种为“南梗46”。每个池塘面积也为10亩, 池塘水深1.2 m, 进排水方便。稻田及池塘水引自当地同一进水渠。在试验期间, 水温变化范围为9.4—30.5℃。试验所用沙塘鳢苗种均为淮安市水生生物研究中心自行繁殖的同批次鱼苗。
1.2 试验设计
试验分为稻田养殖沙塘鳢组(D组)和池塘养殖沙塘鳢组(C组)两种养殖模式。每种养殖模式有3个重复。3个稻田均放养相同密度的沙塘鳢(1 尾/m2), 沙塘鳢的规格为平均(0.986±0.039) g/尾。3个池塘放养相同密度(3 尾/m2)、相同规格[(0.956±0.083) g/尾]的沙塘鳢苗种。养殖稻田和池塘中均放养相同质量的青虾, 放养密度为每亩放养规格为(1.94±0.13) g/尾的抱卵青虾1.5 kg。试验结束后捕捞测定收获情况。试验期间不使用化肥及农药。
1.3 样本采集及处理
分3次进行样本(生物学测量样本和食性样本)采集, 采集时间分别为2022年8月9日、10月8日和12月7日。沙塘鳢和青虾样本采集后, 分别采用游标卡尺和电子天平进行常规生物学测量, 然后冰冻保存。沙塘鳢常规生物学测量包括体长(体长为吻端至尾鳍基部的长度)、体重、口裂宽(鱼嘴巴张开时的水平长度)。青虾测量额剑长(从眼柄基部到额剑前端的距离)及体长(从眼柄基部到尾节末端的长度)。
在食性样本采集时, 每次于3个重复中随机采集50尾, 共计在养殖稻田和池塘各采集150尾沙塘鳢。在对新鲜样本进行解剖之前, 要先观察样本口腔中是否有未被全部吞咽的食物。切开腹腔, 观察胃充塞度(共六级, 0—5), 再仔细切下消化道, 用绳子将两端扎紧, 防止食物外泄。将切下的消化道分别放入已编号且有4%福尔马林溶液的10 mL离心管中保存。食物种类的鉴别是在实验室中进行的。对体型较大、消化程度较低的内含物进行形态学鉴定; 消化程度高的内含物则可以依据咽喉齿(鲫)、鳞片等不易消化的组织进行分类。而体型较小的内含物则借助显微镜来观察鉴定。鉴定工作结束后, 测量部分沙塘鳢消化道中尚未被消化或消化程度低的青虾大小。
1.4 计算公式
成活率(Survival rate, SR, %)=Af /A i×100
相对增重率(Relative weight gain rate, RWG, %)=[(Wt–W0)/W0]×100
湿重特定生长率(Wet weight specific growth rate, SGRw, %/d)=100×(ln Wt–ln W0)/t
摄食率(Feeding rate, FR, %)=(N1/N)×100
出现率(Occurrence rate, OR, %)=(Nm/N)×100
式中, Af为试验末沙塘鳢存活尾数, Ai为试验初沙塘鳢尾数; W0、Wt为试验初和试验末沙塘鳢的体质量(g), N1为消化管有食物鱼数, Nm为胃肠中含有某种食物的鱼数, t为试验时间, N为解剖鱼总数。
1.5 数据分析
分别用Excel 和Origin Pro 8.0软件进行数据处理及相关性分析, 选择R2值高且使用方便的模型为优化模型。本试验数据采用SPSS 19.0软件进行单因素方差分析(One-way ANOVA)及多重比较(Duncan检验)进行分析处理, 以P<0.05作为差异显著水平。用GraphPad Prism 8软件绘图。
2. 结果
2.1 沙塘鳢的存活与生长
池塘和稻田养殖模式下沙塘鳢成活率均在93%以上, 组间无显著差异(表 1; P>0.05)。两种养殖模式显著影响沙塘鳢的终末体长与终末体重和体增重(P<0.05), D组沙塘鳢终末体长、终末体重、体增重(BWG)显著高于C组(P<0.05); D组相对增重率(RWG)及特定生长率(SGRW)高于C组, 但组间无显著差异(P>0.05)。
表 1 养殖模式对沙塘鳢存活率、生长的影响Table 1. The effect of different culture modes on the survival and growth of the O. obscura参数Parameter C组(池塘)
Group C
(Pond culture)D组(稻田)
Group D
(Rice-fish culture)存活率SR (%) 94.58±0.481 93.57±0.353 初始体长IBL (cm) 2.986±0.055 3.02±0.044 终末体长FBL (cm) 14.302±0.043b 14.834±0.030a 终末体重FBW (g) 42.806±0.771b 45.254±0.602a 体增重BWG (g) 41.850±0.755b 44.268±0.569a 相对增重率RWG (%) 4487.154±367.796 4509.706±137.092 湿重特定生长率SGRW (%/d) 3.178±0.065 3.190±0.025 注: 表中各数值(平均值±标准误)后的不同字母表示有显著性差异(P<0.05)Note: Different letters after each value in the table (mean±SE) indicate a significant difference (P<0.05). IBL. initial body length; FBL. final body length; FBW. final body weight; BWG. body weight gain 2.2 沙塘鳢的食物组成
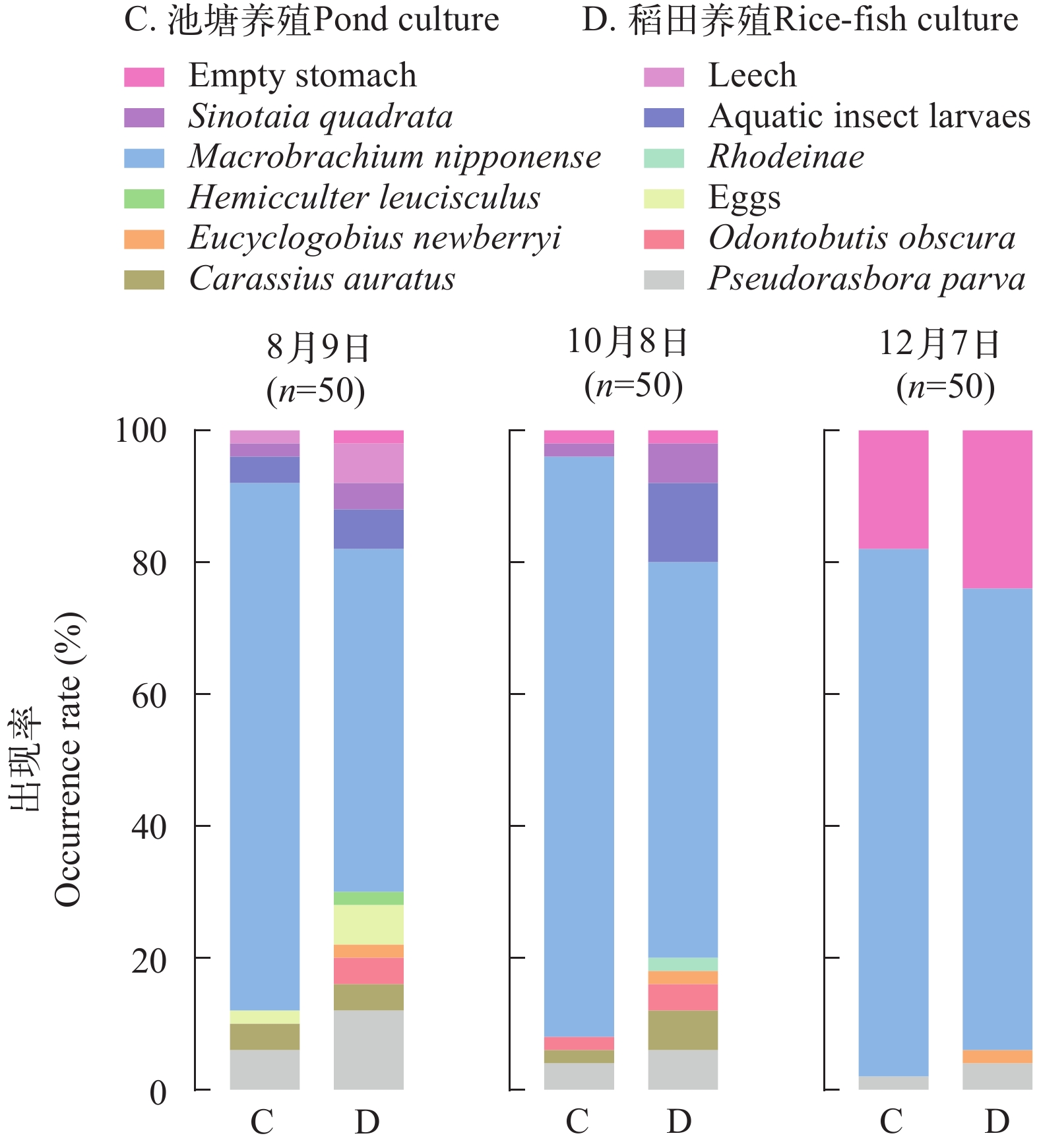
在第1次采样中, C组出现7种饵料生物, 第2次采样新增1种: 沙塘鳢, 第3次无其他种类出现。而D组第1次采样出现10种饵料生物, 第2次新增1种: 鳑鲏, 第3次无新增种类。第3次采样, C组和D组空腹沙塘鳢的条数均增加(图 1)。
![]() 图 1 三次采样期间池塘和稻田中沙塘鳢消化道食物组成情况Figure 1. Food composition of the digestive tract of O. obscura in pond and rice-fish culture during the three sampling periods
图 1 三次采样期间池塘和稻田中沙塘鳢消化道食物组成情况Figure 1. Food composition of the digestive tract of O. obscura in pond and rice-fish culture during the three sampling periodsC组沙塘鳢的摄食率为93.33%, 平均充塞度为3.84。D组沙塘鳢的摄食率为88.00%, 平均充塞度为3.15。C组中除10尾沙塘鳢是空腹, 其余140尾沙塘鳢消化道中均有食物, 且食物种类有8种, 出现率由高到低依次为青虾、麦穗鱼、鲫、水生昆虫幼虫、螺、沙塘鳢、鱼卵和蛭(表 2)。D组中有14尾沙塘鳢是空腹, 其余136尾沙塘鳢消化道中有11种食物, 其出现率由高到低依次为青虾、麦穗鱼、水生昆虫幼虫、鲫、螺、沙塘鳢、虾虎鱼、鱼卵、蛭、䱗和鳑鲏。D组沙塘鳢摄食的食物种类多于C组。两种养殖模式下, 沙塘鳢摄食最多的饵料生物均是青虾, 出现率分别为82.67%(C组)、60.67%(D组)。
表 2 不同养殖模式下沙塘鳢的食物组成Table 2. Food composition of O.obscura under different culture modes食物种类
FoodC组(池塘)
Group C
(Pond culture)D组(稻田)
Group D
(Rice-fish culture)出现次数
Occurrences出现率
Occurrence
rate (%)出现次数
Occurrences出现率
Occurrence
rate (%)鱼类Fishes 麦穗鱼Pseudorasbora parva 6 4.00 11 7.33 鲫Carassius auratus 3 2.00 5 3.33 沙塘鳢Odontobutis obscura 1 0.67 4 2.67 虾虎鱼Eucyclogobius newberryi 0 0 3 2.00 鱼卵Eggs 1 0.67 3 2.00 䱗Hemicculter leucisculus 0 0 1 0.67 鳑鲏Rhodeinae 0 0 1 0.67 虾类Shrimp 青虾Macrobrachium nipponense 124 82.67 91 60.67 水生昆虫幼虫Aquatic insect larvaes 2 1.33 9 6.00 方形环棱螺Sinotaia quadrata 2 1.33 5 3.33 蛭Leech 1 0.67 3 2.00 空腹Empty stomach 10 6.67 14 9.33 2.3 沙塘鳢体长与体重关系
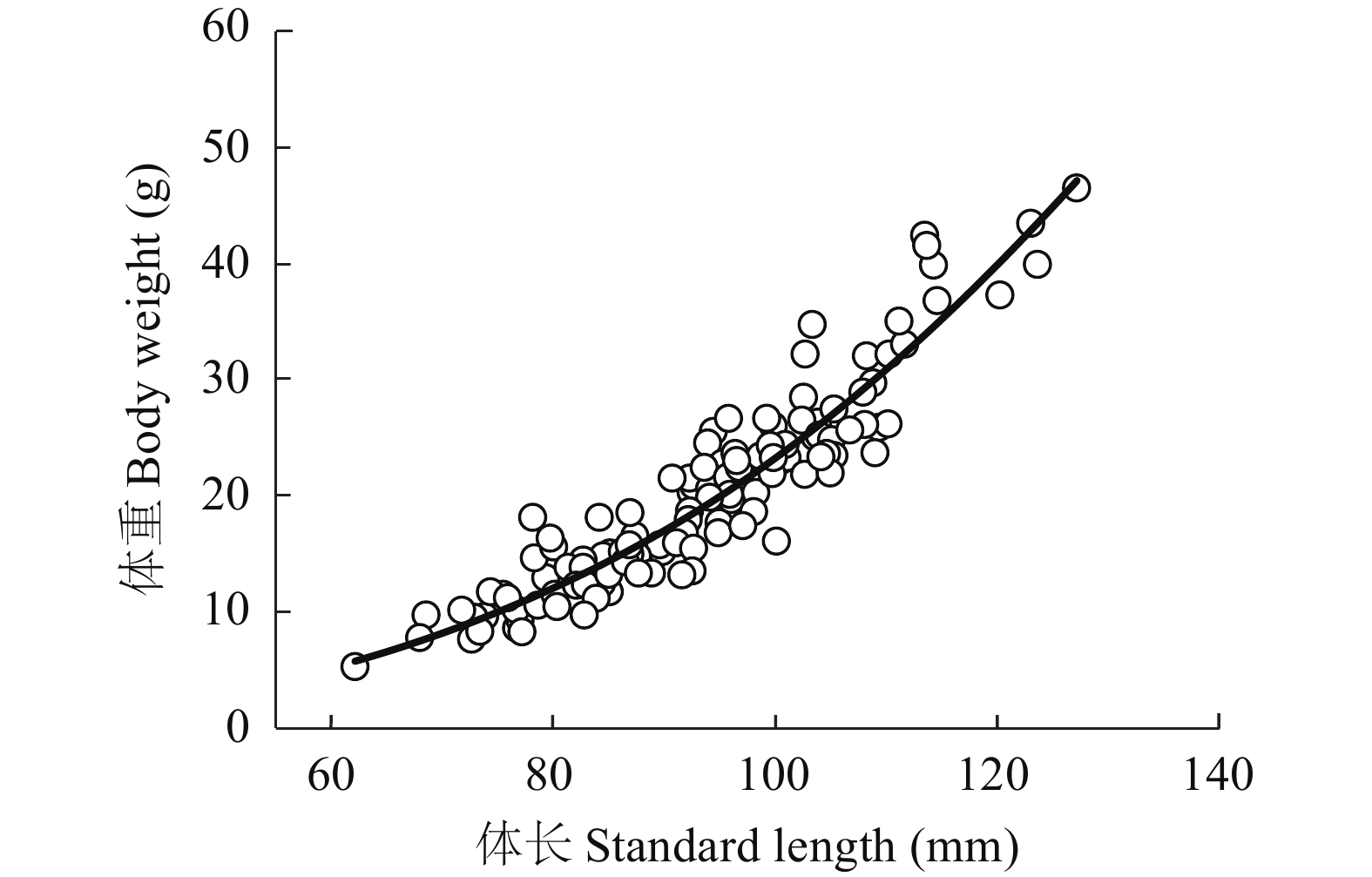
本次试验在稻田和池塘各随机捕获73尾沙塘鳢样本, 经检验, 稻田和池塘两种模式下沙塘鳢体长与体重关系无显著性差异(图 2), 因此对两种养殖模式下其体长与体重关系进行合并拟合, 筛选得出关系模型为y=axb, 即方程W=3.0×10–5L2.9183(n=146; R2=0.879; P<0.05), W为体重, L为体长。
![]() 图 2 沙塘鳢体长与体重关系Figure 2. The relationship between body length and body weight of O. obscura
图 2 沙塘鳢体长与体重关系Figure 2. The relationship between body length and body weight of O. obscura2.4 沙塘鳢、青虾生长参数关系
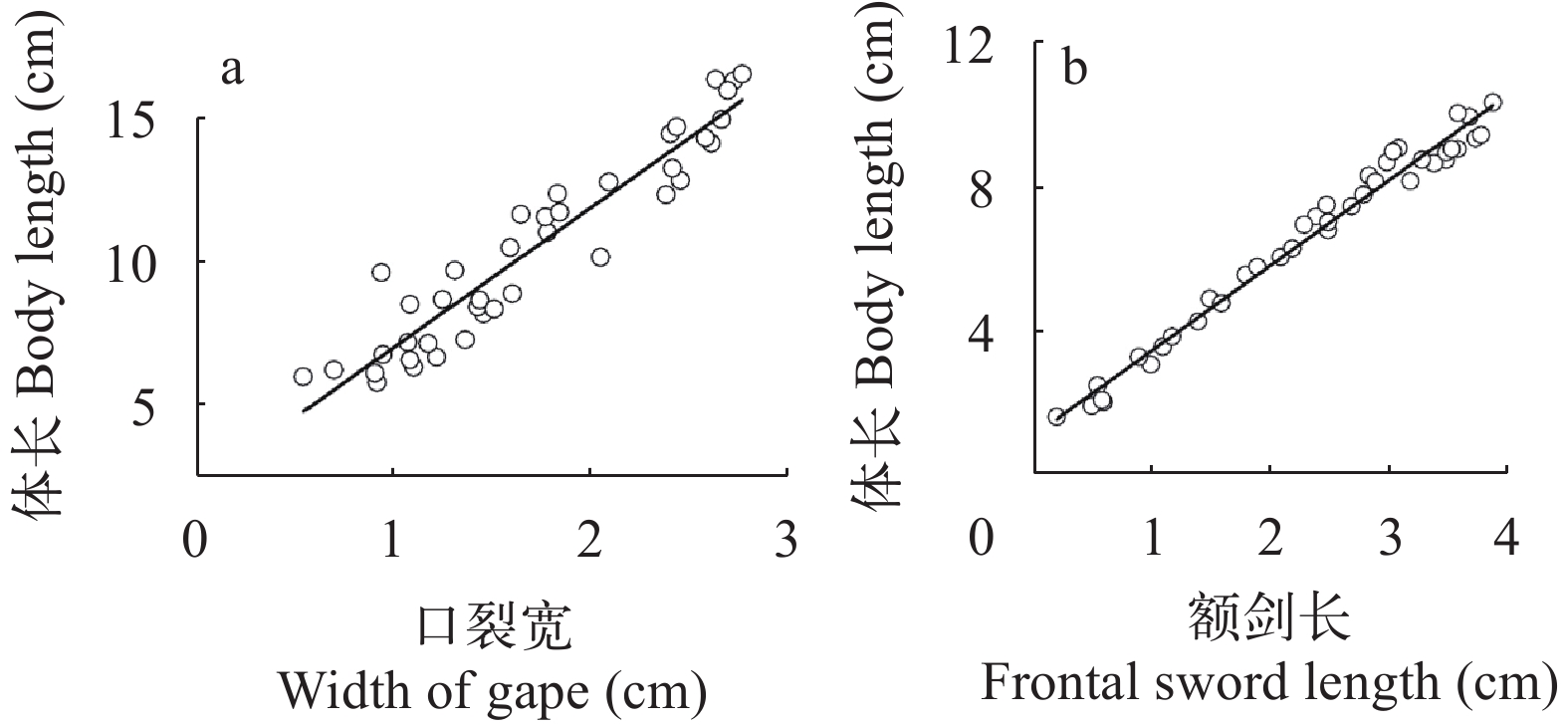
为进一步研究沙塘鳢与其主要食物组成青虾生长参数关系, 对两者生长指标进行相关性分析。本次试验沙塘鳢、青虾及沙塘鳢与青虾之间的关系均以模型y=a+bx拟合较好, 且池塘与稻田两种模式下, 差异不显著, 所以以下均为合并后的方程。本次测量的40尾(稻田和池塘各20尾, 3个重复中随机捕获)沙塘鳢样本口裂宽幅度为0.56—2.70 cm, 体长幅度为5.88—16.02 cm。经拟合, 口裂宽与体长的相关关系式为y=4.865x+2.005(n=40; R2=0.901, P<0.05), 呈线性相关(图 3a)。本次测量的40尾青虾(来自测量的40尾沙塘鳢的消化道中)样本额剑长幅度为0.21—3.95 cm, 体长幅度为1.62—10.45 cm。经拟合, 青虾额剑长与体长的相关关系式为y=2.370x+1.082(n=40; R2=0.980; P<0.05), 两者呈线性相关(图 3b)。
![]() 图 3 沙塘鳢口裂宽与体长的相关关系(a)及青虾额剑长与体长的相关关系(b)Figure 3. Correlation between the width of gape and body length of O. obscura (a) and the Correlation between frontal sword length and body length of M. nipponense (b)
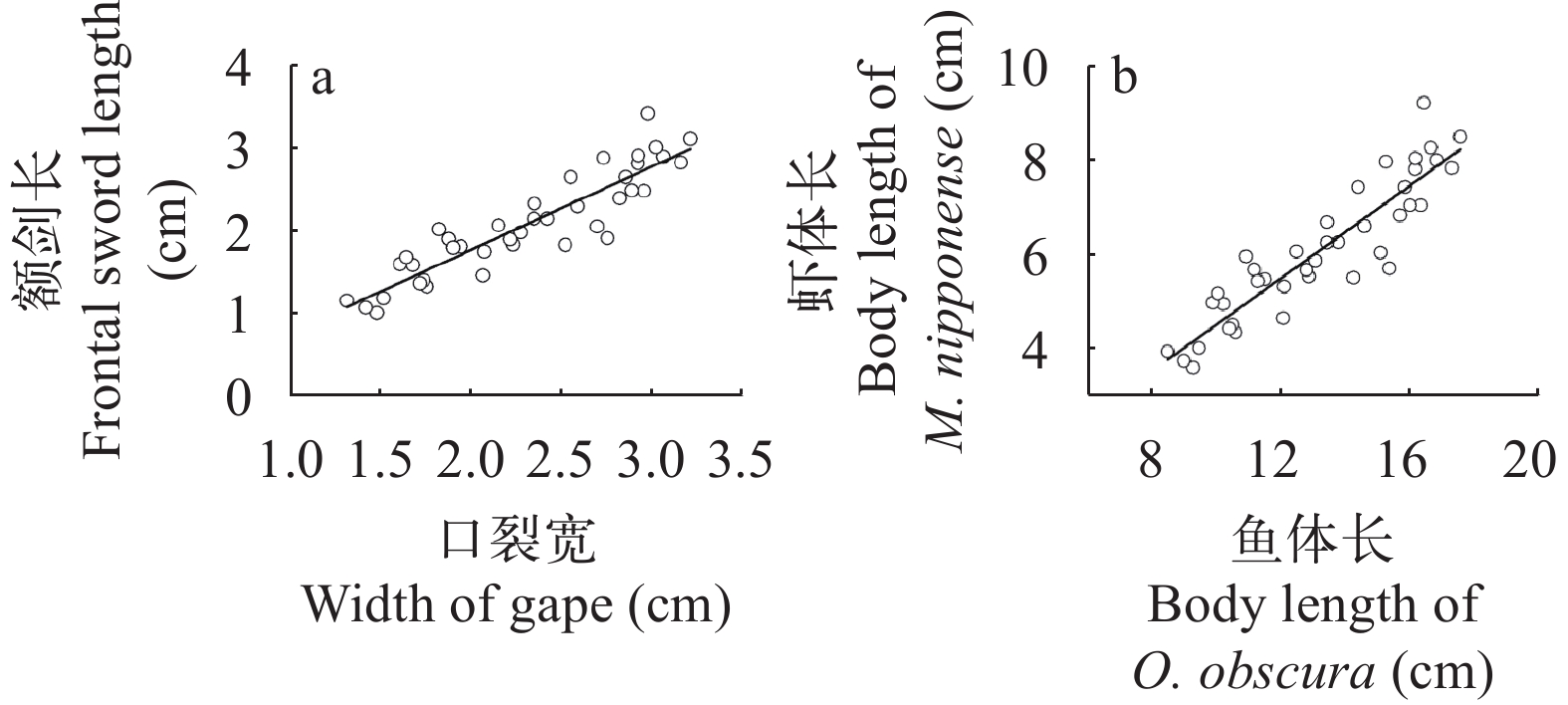
图 3 沙塘鳢口裂宽与体长的相关关系(a)及青虾额剑长与体长的相关关系(b)Figure 3. Correlation between the width of gape and body length of O. obscura (a) and the Correlation between frontal sword length and body length of M. nipponense (b)经拟合, 本次测量的沙塘鳢食性样本口裂宽与其消化道中的青虾额剑长呈现线性相关, 相关关系式为y=1.012x–0.230(n=40; R2=0.828; P<0.05; 图 4a)。根据前期分析得出的相关关系式, 拟合出沙塘鳢体长与其摄食的青虾体长呈现线性关系, 关系为y=0.493x–0.452(n=40; R2=0.828; P<0.05; 图 4b)。
![]() 图 4 沙塘鳢口裂宽与青虾额剑长的关系(a)和沙塘鳢体长与青虾体长的关系(b)Figure 4. Relationship between the width of gape of O. obscura and the length of the frontal sword of M. nipponense (a) and the relationship between the body length of O. obscura and the body length of M. nipponense (b)
图 4 沙塘鳢口裂宽与青虾额剑长的关系(a)和沙塘鳢体长与青虾体长的关系(b)Figure 4. Relationship between the width of gape of O. obscura and the length of the frontal sword of M. nipponense (a) and the relationship between the body length of O. obscura and the body length of M. nipponense (b)3. 讨论
3.1 养殖模式对沙塘鳢存活、生长及食物组成的影响
稻田养殖是我国生态循环农业经济的主要模式之一, “十二五”和“十三五”期间, 得到了长足发展, 在促进乡村振兴、脱贫攻坚和渔业高质量发展等方面发挥了重要作用。在稻田养殖模式中, 养殖鱼类与稻田互利共生的关系, 使其成为一种生态化、绿色化、高效化的养殖模式[3]。本研究通过对比稻田和池塘两种养殖模式发现, 养殖模式对沙塘鳢的存活没有显著影响, 但稻田组沙塘鳢的生长指标(RWG和SGRW)均高于池塘组, 这表明稻田养殖沙塘鳢日均体重增长更快。这一结论在其他鱼类的研究中也有报道。Jeanrenaud等[11]发现稻鱼养殖系统中尼罗罗非鱼(Oreochromis niloticus)平均体重和特定生长率均高于单一养鱼组。吕敏等[12]研究表明稻田养殖模式下瓦氏黄颡鱼(Pseudobagrus vachelli)的体长、体质量、肥满度和特定生长率均显著高于池塘养殖瓦氏黄颡鱼。以上研究结果均表明稻田养殖提高了养殖鱼类的生长速度。这可能是由于稻田中食物资源更加丰富, 沙塘鳢可摄食种类增加导致的。
在两种养殖模式下, 沙塘鳢消化道中出现率最高的均是青虾, 其次是麦穗鱼。这与朱邦科等[13]对保安湖沙塘鳢食性研究结果相似, 保安湖沙塘鳢消化道的食物组成中虾类及小型底栖鱼类的出现率最高。在3次采样中, 稻田养殖沙塘鳢消化道中的饵料生物种类均多于池塘养殖沙塘鳢, 且除青虾外, 稻田养殖沙塘鳢消化道中饵料生物出现次数也多于池塘养殖沙塘鳢, 这表明稻田具有更丰富的食物资源。稻田可以提供养殖生物优越的生活条件, 一般情况下, 鱼类捕食遵循最优觅食理论, 即在最短的处理时间内获得最多的能量[14]。有些鱼会通过选择数量最多的猎物来减少捕获猎物的成本(能量和时间)[15]。相较于池塘, 稻田中丰富的食物资源, 增加了沙塘鳢与饵料生物的相遇率, 减少了沙塘鳢捕食所需的能量消耗, 使得沙塘鳢能够获得更好的生长。
在本试验中, 池塘养殖沙塘鳢的摄食率和胃充塞度都高于稻田养殖沙塘鳢, 但稻田养殖沙塘鳢的终末体长、体增重等指标高于池塘养殖沙塘鳢, 说明稻田环境中的某些因素有利于沙塘鳢的生长。有研究表明稻田养殖模式有利于改善水质, 提高稻田生态系统的稳定性, 从而有利于养殖生物的生活[16]。Li等[17]研究表明稻渔共作模式下, 稻田中总氮、氨氮、总磷等含量均降低。Feng等[18]研究显示黄颡鱼(Pelteobagrus fulvidraco)稻田养殖系统水体中总氮、氨氮、硝态氮、亚硝酸盐-n、总磷和正磷酸盐的含量都低于单一养鱼系统。
3.2 沙塘鳢体长与体重的关系及沙塘鳢与青虾的关系
鱼类体长与体重的关系被广泛应用于鱼类生物学方面的研究, 如鱼类生长、性腺发育、摄食率和成熟度等[19]。体长与体重方程W=aLb中, a代表鱼体的肥硕程度, a值越大, 鱼越肥硕, b值代表鱼体的生长状况, b值小于或大于3代表鱼属于异速生长, b值等于3代表鱼属于匀速生长[20]。王英俊等[21]测量了捕获的24种共1002尾鱼的体长与体重, 结果显示24种鱼的体长与体重均呈显著幂函数相关, 分类地位相同(鲈形目)且食性类似的鱼类有10种, 其中b值大于3的有斑尾刺虾虎鱼(Synechogobius ommaturus)与小头栉孔虾虎鱼(Ctenotrypauchen microcephalus), 为正异速生长, 其余8种鱼的b值均小于3, 都为负异速生长。本研究通过拟合沙塘鳢体长与体重的数据, 得出幂指数方程为W=3.0×10–5L2.9183(n=146; R2=0.879; P<0.05)。由b值显著小于匀速生长值3可知, 本试验所测沙塘鳢属于异速生长。这与梁子湖沙塘鳢[22]不同。而a值与b值会受到性别、生存环境、生长发育的影响[23], 所以两者之间的差异可能是环境因素造成的。
大多数肉食性鱼类因其自身口裂的限制或与饵料生物体型的差异而进行有选择性的捕食[24]。例如鳜(Siniperca chuatsi)[25]、大口黑鲈(Micropterus salmoides)[26]等。这些鱼类可摄食的饵料生物范围均会随着口裂、体型的生长而增大。沙塘鳢属于小型底栖肉食性鱼类, 从体型上来讲, 其只能捕食比它自身体型还小的小型水产动物, 例如青虾, 麦穗鱼等。青虾游泳能力较弱[27], 易被捕食, 且是沙塘鳢喜食的一类饵料生物[13]。所以本试验通过青虾来探究沙塘鳢与饵料生物之间的关系。额剑是位于青虾头胸甲前端中央的一个剑状突起, 是青虾区别于其他虾类的重要形态特征[28], 目前其作用机制尚且不明。在本研究中, 青虾额剑长与体长呈线性正相关, 方程为y=2.370x+1.082(n=40; R2=0.980; P<0.05), 说明随着青虾额剑长的生长, 其体长呈匀速增长。沙塘鳢的口裂宽与其体长呈线性正相关, 方程为y=4.865x+2.005(n=40; R²=0.901; P<0.05), 说明随着沙塘鳢口裂宽的增长, 其体长也在匀速增加。青虾额剑长与沙塘鳢口裂宽呈线性正相关, 青虾体长与沙塘鳢体长也呈线性相关, 说明随着沙塘鳢的生长, 其所能摄食的青虾规格也不断增大。保安湖沙塘鳢体长约为其口裂宽的5倍, 可捕食最大猎物体长为其体长的50%—70%[13]。本试验中沙塘鳢体长也约为口裂宽的5倍, 沙塘鳢可摄食的最大青虾长度为其体长的65%(范围上限), 与保安湖沙塘鳢的研究结果相似。
4. 结论
养殖模式对沙塘鳢的存活率没有显著影响, 两种养殖模式下沙塘鳢的存活率均高达93%。稻田养殖沙塘鳢的终末体长、终末体重、体增重、相对增重率及特定生长率均高于池塘养殖沙塘鳢。虽然池塘沙塘鳢的摄食率高于稻田养殖沙塘鳢, 但稻田养殖沙塘鳢的生长指标较好, 且与池塘养殖模式相比, 稻田环境可以为沙塘鳢提供更丰富的食物资源, 减少养殖成本。因此, 对于沙塘鳢养殖, 稻田养殖模式优于池塘养殖模式。在本试验中, 沙塘鳢体长与体重呈幂函数, b值小于3, 为异速生长; 沙塘鳢口裂宽与其体长、青虾额剑长呈线性正相关, 青虾体长与其额剑长、沙塘鳢体长也呈线性正相关。
-
![]()
图 1 三次采样期间池塘和稻田中沙塘鳢消化道食物组成情况
Figure 1. Food composition of the digestive tract of O. obscura in pond and rice-fish culture during the three sampling periods
![]()
图 2 沙塘鳢体长与体重关系
Figure 2. The relationship between body length and body weight of O. obscura
![]()
图 3 沙塘鳢口裂宽与体长的相关关系(a)及青虾额剑长与体长的相关关系(b)
Figure 3. Correlation between the width of gape and body length of O. obscura (a) and the Correlation between frontal sword length and body length of M. nipponense (b)
![]()
图 4 沙塘鳢口裂宽与青虾额剑长的关系(a)和沙塘鳢体长与青虾体长的关系(b)
Figure 4. Relationship between the width of gape of O. obscura and the length of the frontal sword of M. nipponense (a) and the relationship between the body length of O. obscura and the body length of M. nipponense (b)
表 1 养殖模式对沙塘鳢存活率、生长的影响
Table 1 The effect of different culture modes on the survival and growth of the O. obscura
参数Parameter C组(池塘)
Group C
(Pond culture)D组(稻田)
Group D
(Rice-fish culture)存活率SR (%) 94.58±0.481 93.57±0.353 初始体长IBL (cm) 2.986±0.055 3.02±0.044 终末体长FBL (cm) 14.302±0.043b 14.834±0.030a 终末体重FBW (g) 42.806±0.771b 45.254±0.602a 体增重BWG (g) 41.850±0.755b 44.268±0.569a 相对增重率RWG (%) 4487.154±367.796 4509.706±137.092 湿重特定生长率SGRW (%/d) 3.178±0.065 3.190±0.025 注: 表中各数值(平均值±标准误)后的不同字母表示有显著性差异(P<0.05)Note: Different letters after each value in the table (mean±SE) indicate a significant difference (P<0.05). IBL. initial body length; FBL. final body length; FBW. final body weight; BWG. body weight gain  下载: 导出CSV
下载: 导出CSV
表 2 不同养殖模式下沙塘鳢的食物组成
Table 2 Food composition of O.obscura under different culture modes
食物种类
FoodC组(池塘)
Group C
(Pond culture)D组(稻田)
Group D
(Rice-fish culture)出现次数
Occurrences出现率
Occurrence
rate (%)出现次数
Occurrences出现率
Occurrence
rate (%)鱼类Fishes 麦穗鱼Pseudorasbora parva 6 4.00 11 7.33 鲫Carassius auratus 3 2.00 5 3.33 沙塘鳢Odontobutis obscura 1 0.67 4 2.67 虾虎鱼Eucyclogobius newberryi 0 0 3 2.00 鱼卵Eggs 1 0.67 3 2.00 䱗Hemicculter leucisculus 0 0 1 0.67 鳑鲏Rhodeinae 0 0 1 0.67 虾类Shrimp 青虾Macrobrachium nipponense 124 82.67 91 60.67 水生昆虫幼虫Aquatic insect larvaes 2 1.33 9 6.00 方形环棱螺Sinotaia quadrata 2 1.33 5 3.33 蛭Leech 1 0.67 3 2.00 空腹Empty stomach 10 6.67 14 9.33
下载: 导出CSV
-
[1] 高鸣, 陈洁, 姚志. 中国淡水养殖业绿色发展: 提质增效与未来路径 [J]. 华中农业大学学报(自然科学版), 2022, 41(3): 96-106. Gao M, Chen J, Yao Z. Green development of freshwater aquaculture in China: improving quality and increasing efficiency and the future path [J]. Journal of Huazhong Agricultural University (Natural Science Edition), 2022, 41(3): 96-106.
[2] 王强盛, 王晓莹, 杭玉浩, 等. 稻田综合种养结合模式及生态效应 [J]. 中国农学通报, 2019, 35(8): 46-51. doi: 10.11924/j.issn.1000-6850.casb18040052 Wang Q S, Wang X Y, Hang Y H, et al. Combination modes and ecological effects of planting-breeding ecosystem in rice field [J]. Chinese Agricultural Science Bulletin, 2019, 35(8): 46-51. doi: 10.11924/j.issn.1000-6850.casb18040052
[3] Li F F, Gao J C, Xu Y, et al. Biodiversity and sustainability of the integrated rice-fish system in Hani terraces, Yunnan province, China [J]. Aquaculture Reports, 2021(20): 100763.
[4] 步洪凤, 梁玉刚, 方宝华. 稻田综合种养主要模式及其研究进展 [J]. 杂交水稻, 2023, 38(1): 10-19. doi: 10.16267/j.cnki.1005-3956.20220409.170 Bu H F, Liang Y G, Fang B H. Main models and research progress of planting-breeding ecosystem in rice field [J]. Hybrid Rice, 2023, 38(1): 10-19. doi: 10.16267/j.cnki.1005-3956.20220409.170
[5] Liang X F, Lin X T, Li S Q, et al. Impact of environmental and innate factors on the food habit of Chinese perch Siniperca chuatsi (Basilewsky) (Percichthyidae) [J]. Aquaculture Research, 2008, 39(2): 150-157. doi: 10.1111/j.1365-2109.2007.01870.x
[6] Krebs J M, Turingan R G. Intraspecific variation in gape–prey size relationships and feeding success during early ontogeny in red drum, Sciaenops ocellatus [J]. Environmental Biology of Fishes, 2003, 66(1): 75-84. doi: 10.1023/A:1023290226801
[7] 胡亚丽, 朱晓平, 尹绍武, 等. 河川沙塘鳢的含肉率及肌肉营养价值分析 [J]. 江苏农业科学, 2012, 40(7): 290-293. doi: 10.3969/j.issn.1002-1302.2012.07.111 Hu Y L, Zhu X P, Yin S W, et al. Analysis of meat content and muscle nutritional value of Odontobutis potamophilus [J]. Jiangsu Agricultural Sciences, 2012, 40(7): 290-293. doi: 10.3969/j.issn.1002-1302.2012.07.111
[8] 赵彦华, 刘洪岩, 孙梦玲, 等. 养殖沙塘鳢烂鳃症病原研究及最适治疗药物的筛选 [J]. 江苏农业科学, 2018, 46(9): 171-173. doi: 10.15889/j.issn.1002-1302.2018.09.040 Zhao Y H, Liu H Y, Sun M L, et al. Study on pathogens of gill disease of Odontobutis obscura in breeding ponds and screening of the most suitable therapeutic drugs [J]. Jiangsu Agricultural Sciences, 2018, 46(9): 171-173. doi: 10.15889/j.issn.1002-1302.2018.09.040
[9] 郑国平. 沙塘鳢池塘养殖与管理技术 [J]. 农技服务, 2012, 29(1): 67-68. doi: 10.3969/j.issn.1004-8421.2012.01.050 Zheng G P. Pond culture and management techniques of Odontobutis sinensis [J]. Agricultural Technology Service, 2012, 29(1): 67-68. doi: 10.3969/j.issn.1004-8421.2012.01.050
[10] 蒋刚, 戴颖, 张亚, 等. 温度对日本沼虾幼体发育及成体繁殖性能的影响 [J]. 上海海洋大学学报, 2022, 31(2): 365-372. Jiang G, Dai Y, Zhang Y, et al. Effects of temperature on the growth and reproduction performance of Macrobrachium nipponense from zoea to postlarvae [J]. Journal of Shanghai Ocean University, 2022, 31(2): 365-372.
[11] Jean-Renaud A, Emmanuel A N, Yves B K, et al. Evaluation of performance of Nile tilapia (Oreochromis niloticus) crop in rice-fish ponds [J]. Journal of Entomology and Zoology Studies, 2016, 4(1): 91-97.
[12] 吕敏, 甘晖, 陈田聪, 等. 瓦氏黄颡鱼在稻田和池塘养殖中的生长性能和肌肉品质比较 [J]. 水产学杂志, 2022, 35(1): 75-81. doi: 10.3969/j.issn.1005-3832.2022.01.013 Lü M, Gan H, Chen T C, et al. Comparison of growth performance and muscle quality of yellow catfish (Pseudobagrus vachelli) cultured in rice fields and ponds [J]. Chinese Journal of Fisheries, 2022, 35(1): 75-81. doi: 10.3969/j.issn.1005-3832.2022.01.013
[13] 朱邦科, 谢从新, 王明学, 等. 保安湖沙塘鳢的食性、繁殖、年龄及生长的研究 [J]. 水生生物学报, 1999, 23(4): 316-323. doi: 10.3321/j.issn:1000-3207.1999.04.004 Zhu B K, Xie C X, Wang M X, et al. A study on feeding, reproduction, age and growth of dark sleeper Odontobutis obscura in Bao’an Lake [J]. Acta Hydrobiologica Sinica, 1999, 23(4): 316-323. doi: 10.3321/j.issn:1000-3207.1999.04.004
[14] Turesson H, Persson A, Brönmark C. Prey size selection in piscivorous pikeperch (Stizostedion lucioperca) includes active prey choice [J]. Ecology of Freshwater Fish, 2002, 11(4): 223-233. doi: 10.1034/j.1600-0633.2002.00019.x
[15] Makrakis M C, Nakatani K, Bialetzki A, et al. Relationship between gape size and feeding selectivity of fish larvae from a Neotropical reservoir [J]. Journal of Fish Biology, 2008, 72(7): 1690-1707. doi: 10.1111/j.1095-8649.2008.01845.x
[16] 赵翔刚, 罗衡, 刘其根, 等. 稻田养殖沙塘鳢对稻田水体及底泥微生物群落结构及多样性的影响 [J]. 淡水渔业, 2017, 47(4): 8-14. doi: 10.3969/j.issn.1000-6907.2017.04.003 Zhao X G, Luo H, Liu Q G, et al. Influence of the cultured Odontobutis obscurus to the microbial community structure and diversity in rice-fish system [J]. Freshwater Fisheries, 2017, 47(4): 8-14. doi: 10.3969/j.issn.1000-6907.2017.04.003
[17] Li F B, Sun Z P, Qi H Y, et al. Effects of rice-fish co-culture on oxygen consumption in intensive aquaculture pond [J]. Rice Science, 2019, 26(1): 50-59. doi: 10.1016/j.rsci.2018.12.004
[18] Feng J F, Li F B, Zhou X Y, et al. Nutrient removal ability and economical benefit of a rice-fish co-culture system in aquaculture pond [J]. Ecological Engineering, 2016(94): 315-319.
[19] Amouei F, Valinassab T, Haitov A. Age determination and morphological study using otoliths in Cyprinus carpio Linnaeus, 1758 in the Southern Caspian Sea [J]. Iranian Journal of Fisheries Sciences, 2013, 12(4): 759-769.
[20] 邢迎春, 赵亚辉, 张洁, 等. 北京地区宽鳍鱲的生长及食性 [J]. 动物学报, 2007, 53(6): 982-993. doi: 10.3969/j.issn.1674-5507.2007.06.007 Xing Y C, Zhao Y H, Zhang J, et al. Growth and diets of Zacco platypus distributed in Beijing [J]. Acta Zoologica Sinica, 2007, 53(6): 982-993. doi: 10.3969/j.issn.1674-5507.2007.06.007
[21] 王英俊, 吴莹莹, 邹琰, 等. 青岛沿海24种鱼类耳石秤量特征与鱼体大小关系 [J]. 海洋科学, 2018, 42(8): 48-55. doi: 10.11759/hykx20180428001 Wang Y J, Wu Y Y, Zou Y, et al. Relationship between fish size and otolith measurements for fish species caught in the coast of Qingdao, China [J]. Marine Sciences, 2018, 42(8): 48-55. doi: 10.11759/hykx20180428001
[22] 曹晋飞. 梁子湖中华沙塘鳢年龄生长、胚胎发育和消化道组织学的研究 [D]. 武汉: 华中农业大学, 2014: 11-16. Cao J F. Studies on the age, growth, embryonic development and histology of digestive tract of Odontobutis sinensis [D]. Wuhan: Huazhong Agricultural University, 2014: 11-16.
[23] 王普泽, 宋聃, 张尹哲, 等. 博斯腾湖鱼类资源组成、体长与体重关系及生长状况 [J]. 生物资源, 2020, 42(2): 181-187. Wang P Z, Song D, Zhang Y Z, et al. Resource composition, length-weight relationship and condition factor of fishes [J]. Biotic Resources, 2020, 42(2): 181-187.
[24] 任艳, 马琴, 刘怡然, 等. 河川沙塘鳢对猎物种类和大小选择性研究 [J]. 水生生物学报, 2022, 46(6): 865-871. doi: 10.7541/2022.2021.0374 Ren Y, Ma Q, Liu Y R, et al. Prey species and size selection of Odontobutis potamophila [J]. Acta Hydrobiologica Sinica, 2022, 46(6): 865-871. doi: 10.7541/2022.2021.0374
[25] Li W, Zhang T, Ye S et al. Feeding habits and predator-prey size relationships of mandarin fish Siniperca chuatsi (Basilewsky) in a shallow lake, central China [J]. Journal of Applied Ichthyology, 2013, 29(1): 56-63. doi: 10.1111/j.1439-0426.2012.02044.x
[26] Einfalt L M, Parkos J J, Wahl D H. Effect of predator size and prey characteristics on piscivory of juvenile largemouth bass [J]. Transactions of the American Fisheries Society, 2015, 144(4): 682-692. doi: 10.1080/00028487.2015.1027002
[27] 傅洪拓, 蒋速飞. 青虾新品种池塘主养技术 [J]. 科学养鱼, 2020(4): 26-27. doi: 10.3969/j.issn.1004-843X.2020.04.013 Fu H T, Jiang S F. Pond-culture technique of new freshwater shrimp variety [J]. Scientific Fish Farming, 2020(4): 26-27. doi: 10.3969/j.issn.1004-843X.2020.04.013
[28] 吕丁, 傅洪拓, 乔慧, 等. 青虾种质资源研究与保护进展 [J]. 中国农学通报, 2012, 28(11): 97-102. doi: 10.11924/j.issn.1000-6850.2011-3624 Lü D, Fu H T, Qiao H, et al. Research progress on germplasm resource of Macrobrachium nipponense [J]. Chinese Agricultural Science Bulletin, 2012, 28(11): 97-102. doi: 10.11924/j.issn.1000-6850.2011-3624
-
期刊类型引用(0)
其他类型引用(1)
计量
- 文章访问数: 1360
- HTML全文浏览量: 180
- PDF下载量: 67
- 被引次数: 1