
FLOW VELOCITY ON GROWTH AND BEHAVIOR IN BLACK ROCKFISH (SEBASTES SCHLEGELII)
-
摘要: 文章以许氏平鲉(Sebastes schlegelii)幼鱼为研究对象, 研究了不同流速对许氏平鲉幼鱼生长性能和行为表现的影响。实验共设置4个流速处理组, 分别为0(C组)、0.5 BL/s(L组)、1.5 BL/s(M组)和2.5 BL/s(H组), 实验周期为43d。研究共监测与分析了8个行为指标, 并将其归为运动参数(平均速度和最大加速度)、偏好区域(内、中和外圈累计停留时间)和活动状态(最大轮替次数、活跃性和身体接触)等3类行为状态。结果表明: (1)在整个实验周期中, M组和L组的终体长显著高于C组和H组(P<0.05); M组的终体重、特定生长率和增重率显著高于C组和H组(P<0.05)。(2) 在实验30d后, M组和H组的外圈累计停留时间显著低于C组和L组(P<0.05), 且M组中圈累计停留时间显著高于C组和H组(P<0.05); H组的中圈累计停留时间显著低于其他处理组(P<0.05), 其内圈累计停留时间显著高于其他处理组(P<0.05), 且各处理组间没有显著差异(P>0.05); M组的最大加速度显著高于C组和L组(P<0.05), 且36d后, M组的平均速度显著高于其他所有处理(P<0.05)。综上所述, M组(1.5 BL/s)流速能够促进许氏平鲉的生长, 此外, 流速的变化显著影响着许氏平鲉在不同圈层圈的累计停留时间、区域间轮替次数、活跃性和身体接触等行为特征, 可以将区域偏好及活动状态等指标作为评价许氏平鲉应对不同流速条件的行为响应指标。Abstract: In order to investigate the growth and behavior of juvenile black rockfish (Sebastes schlegelii) under different flow velocities, four treatment groups were set up. The average water flows velocity of the four treatments was maintained at 0, 0.5, 1.5, and 2.5 BL/s, expressed as C, L, M, and H, respectively. The experiment lasted for 43d and eight behavior indicators were analyzed. The results showed the following: (1) The final body lengths in M and L were significantly higher than those in the other two treatment groups (P<0.05); the final weight, specific growth rate and weight gain rate were significantly higher in treatment M than those in treatment C and H (P<0.05); the feed conversion ratio was significantly lower in treatment L than that in treatment H (P<0.05). (2) After 30d of the experiment, the accumulated residence time of the outer-circle area was significantly lower in both the M and H treatment groups compared with C and L treatment groups (P<0.05); the accumulated residence time in the middle-circle area was significantly higher in M and L treatment groups (P<0.05); the accumulated residence time of inner-circle area in treatment H was significantly higher than that in other treatment groups (P<0.05); the maximum accelerations of M were significantly higher than those of C and L treatment (P<0.05), after 36d of the experiment, the average velocity of the M treatment was significantly higher than that in the other treatment groups (P<0.05). In conclusion, moderate swimming training significantly improved the growth performance of juvenile black rockfish, and the optimal flow velocity occurred at 1.5 BL/s under the present experimental conditions. Flow velocity significantly influenced the behavioral characteristics of juvenile black rockfish. Behavioral characteristics, such as regional preference and activity status, could be used to assess the respond of juvenile black rockfish to various water flow velocities.
-
Keywords:
- Flow velocity /
- Growth performance /
- Behavior /
- Sebastes schlegelii
-
在集约化养殖过程中, 流速最早是出于促进集污, 降低养殖系统清洁压力的目的而提出[1, 2]。近年来, 越来越多的研究表明, 流速也是影响鱼类生长的重要环境因子之一, 适当的流速可以改善鱼类的生长性能[3—5], 但流速对鱼类生长性能的影响随鱼种的不同而不同[6—8]。Li等[9]研究了不同流速对大菱鲆(Scophthalmus maximus)生长性能的影响, 结果显示, 与其他流速组相比, 大菱鲆在0.9 BL/s的流速下摄食量增加, 并获得了更高的特定生长率。Liu等[10]通过研究不同流速对齐口裂腹鱼(Schizothorax prenanti)生长的影响, 结果发现当流速为2.0 BL/s时, 齐口裂腹鱼的特定生长率和饲料系数显著高于其他处理组。Chen等[11]对大口黑鲈(Micropterus salmoides)幼鱼在不同流速条件下的生长进行了研究, 结果表明在4.0 BL/s时, 大口黑鲈的饲料系数及脂质利用率上升, 其增重率和特定生长率显著高于其他流速处理组。
鱼类具有趋流性, 能够通过侧线感知水流, 并根据水流调整自身的游泳行为以适应水流的变化[12]。对养殖条件下鱼类行为的研究表明, 在适宜的流速条件下, 鱼群会在养殖池中均匀分布, 表现出明显的集群行为, 这对减少鱼群的结构变换和个体交互压力有积极的促进作用[13]。而在较高的流速条件下, 鱼必须专注游泳以应对高流速的冲击, 且需要分配更多的能量用于稳定游泳活动, 这可能会使鱼处于较高的压力水平[14]; 此外, 高流速也会增加鱼与障碍物或其他个体的碰撞概率, 导致新的鳍损伤, 从而影响其生长和福利状况[15, 16]。这一方面说明, 通过鱼类的行为表现能够反映鱼类是否处于不利的环境条件, 另一方面, 随着技术的发展和鱼类福利水平要求的提高, 对养殖过程中鱼类行为的研究可能是改善鱼类性能的一种有效方式, 对养殖的有效性和可持续性至关重要[17—19]。
许氏平鲉(Sebastes schlegelii), 又名黑鲪, 属鲉形目(Scorpaeniformes), 鲉科(Scorpaenidae)平鲉属(Sebastes), 主要分布于中国、韩国和日本, 在我国主要分布于东海、黄海和渤海[20]。由于抗病力较强和生长速度较快, 许氏平鲉已成为近年来中国北方重要的增养殖鱼种之一。但目前关于鱼类在不同流速条件下的研究大都集中在鲑鳟鱼类[21—23], 对养殖条件下许氏平鲉应对不同流速的生长和行为的综合研究尚属空白。本研究以许氏平鲉幼鱼为对象, 探讨了不同流速条件下许氏平鲉幼鱼的生长性能和行为表现规律, 并根据以上结果, 确定了许氏平鲉养殖条件下的最佳流速条件, 以促进其达到最佳生长性能。研究结果能够为许氏平鲉集约化养殖适宜流速的设置提供数据支撑, 为研究不同流速条件下鱼类行为响应特点提供理论依据。
1. 材料与方法
1.1 实验动物
实验鱼为许氏平鲉幼鱼, 从大连天正实业有限公司(中国辽宁)获得。实验在大连海洋大学设施渔业教育部重点实验室进行。实验开始前, 幼鱼在聚乙烯塑料桶中暂养2周, 暂养期间的水温控制在(19.0±0.5)℃, pH为8.00±0.30, 溶解氧>6.5 mg/L, 氨氮<0.02 mg/L, 亚硝态氮<0.05 mg/L, 光暗周期为12L﹕12D。每天8:30和16:30各饱食投喂1次浮性商业饲料(粗蛋白质≥44%, 粗脂肪≥5%)。
1.2 实验装置
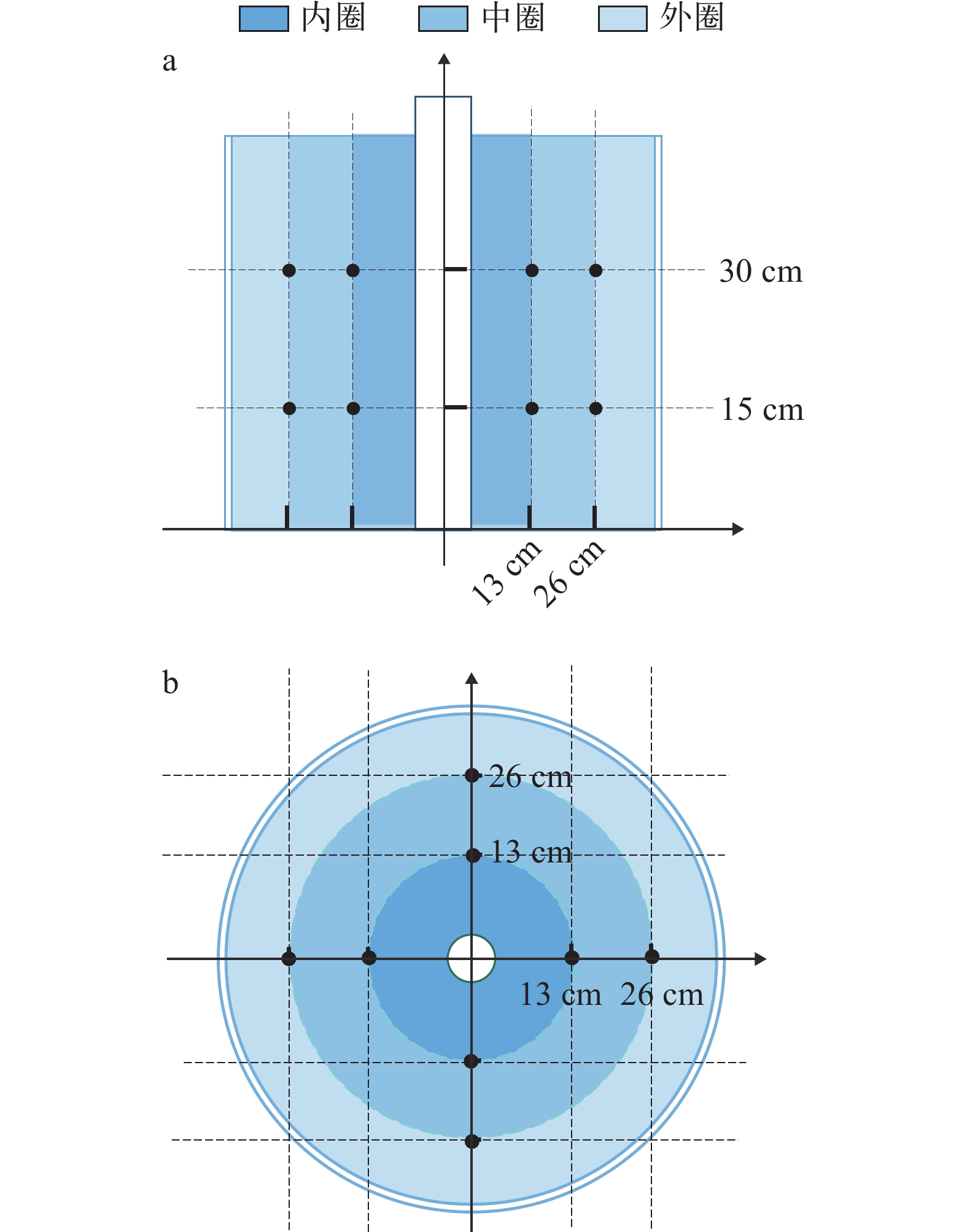
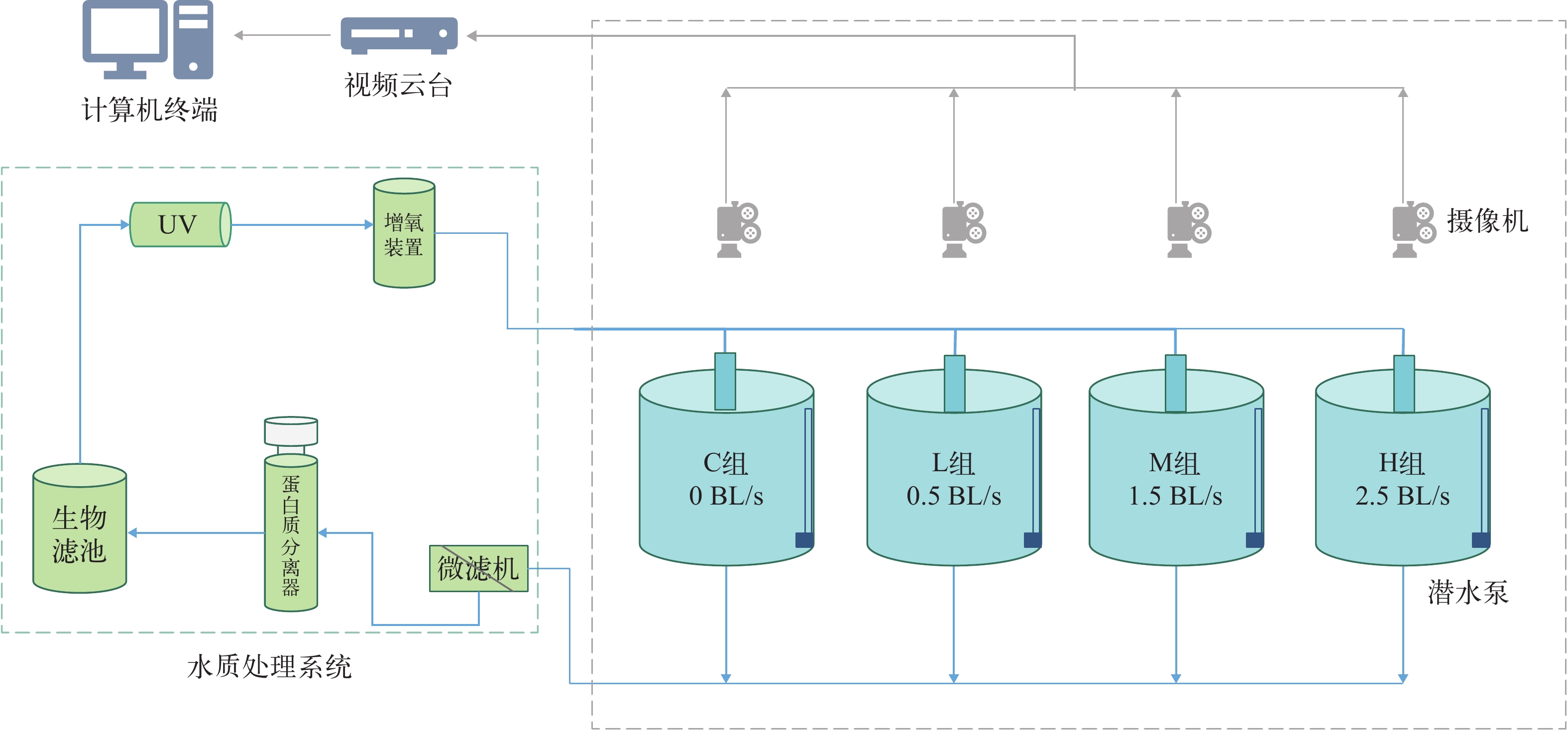
实验在改装过的可调节流速的养殖桶内(r=40 cm, h=80 cm; 有效水体约为200 L)进行, 装置动力由潜水泵提供, 养殖桶侧壁立有一根垂直于桶底的出水管, 水管侧壁有等距出水孔, 潜水泵带动水流从出水孔流出, 在桶内形成稳定环流。通过调节水泵的功率, 来控制实验桶内的水流速度, 直至达到实验预设流速(图 1)。用LS300-A型便携式流速流量仪(南京卓玛机电有限公司)对桶内的流速进行测量: 分别在两个相互垂直的截面上选取了16个点(图 2), 并计算这些点的流速, 用平均值来表示每个养殖桶内的流速, 每个点测量3次[11]。在实验的开始和结束时分别进行了无鱼测量, 每周检查两次水流速度并进行调整, 使水流保持在设定流速范围内。
![]() 图 2 采样点及分析区示意图a. 养殖桶侧视图; b. 养殖桶俯视图Figure 2. Schematic diagram of measurement points and analysis areasa. lateral view of rearing tank; b. top view of rearing tank
图 2 采样点及分析区示意图a. 养殖桶侧视图; b. 养殖桶俯视图Figure 2. Schematic diagram of measurement points and analysis areasa. lateral view of rearing tank; b. top view of rearing tank1.3 实验设计
本实验设定对照组与3个流速处理组, 分别为0(C组)、0.5 BL/s(L组)、1.5 BL/s(M组)和2.5 BL/s(H组), 对应的流速分别为0、4.7、14.2和23.7 cm/s, 每组设置3个平行, 实验共进行43d。选择初始体长(9.57±0.09) cm, 初始体重(21.73±0.16) g的许氏平鲉共192尾, 随机分配到12个养殖桶中, 每个养殖桶内有16尾鱼。在养殖桶上方1 m处架设高清摄像机(HIKDS-2CD3T35D-I5, 海康威视), 用于拍摄和记录实验期间幼鱼的行为。
在实验期间, 各养殖桶的水质参数、光温条件、投喂量和投喂时间与暂养期间保持一致。饲喂期间各组水流保持静止状态, 喂食20min后采用虹吸方法吸出残饵, 收集在筛网滤袋中, 然后置于80℃的烘箱中烘干至恒重后, 用电子天秤称量(精确至0.01 mg), 计算残饵干重。
1.4 生长数据采集
于实验开始的1d、8d、15d、22d、29d、36d和43d随机捞取各处理组3尾鱼, 在浓度为60 mg/L的丁香酚溶液中麻醉, 称量每尾鱼的湿重、体长并记录, 然后依次放回。生长性能相关指标计算方法如下:
存活率(Survival rate, SR, %)=100×最终尾数/初始尾数
特定生长率(Specific growth rate, SGR, %/d)=100×[(lnW2–lnW1)]/t
增重率(Weight gain rate, WGR, %)=(W2–W1)/W1
饲料系数(Feed conversion ratio, FCR)=F/(W2–W1)
式中, W1为初始体重(Initial weight, g); W2为终体重(Final weight, g); t为实验天数; F为摄食量。
1.5 行为数据采集及分析
于实验开始的1d、8d、15d、22d、29d、36d和43d早上8:15(生长采样前), 采集投喂前15min的视频, 并从中截取10min。利用行为学分析软件(Noldus Ethovision XT 12.0)对行为视频进行分析, 基于本实验设计情况, 选择使用动态减影法进行目标识别。共选取8个行为参数来描述不同流速下许氏平鲉的行为, 分别为: 平均速度(Average velocity, cm/s)、最大加速度(The maximum acceleration, cm/s2)、分析区累计停留时间(Accumulated residence time in analysis area, %)、最大轮替次数(Maximum zone alternation times, 次)、活跃性(Activity, %)和身体接触(Physical contact, 次)。其中, 平均速度通过许氏平鲉的移动距离除以观测时间得到; 最大加速度是通过计算许氏平鲉移动时的速度差值除以时间差值得到; 分析区累计停留时间是指许氏平鲉在不同选定区域内的总停留时间; 最大轮替次数是指在观测时间内许氏平鲉在选定的分析区(圈层)中访问的最大次数; 活跃性指的是许氏平鲉的活动情况, 通过对比视频图像像素变化的百分比进行计算; 身体接触是指许氏平鲉在活动过程中身体接触的总次数; 此外, 如图 2所示, 为了分析不同流速条件下幼鱼的区域停留偏好(分析区停留时间), 将视频视野划分为内圈、中圈和外圈3个分析区, 半径依次为13、26和40 cm。相关行为指标的计算公式如下:
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
1.6 数据处理
所有统计分析均采用SPSS 25.0软件进行。统计数据均用平均值±标误差(mean±SE)表示, 使用单因素方差分析(One-way ANOVA)和Tukey事后检验进行分析。显著性水平为P<0.05, 所有表格均使用Excel制作。
2. 结果
2.1 流速对许氏平鲉生长的影响
如表 1所示, 所有处理组中的SR均为100%。H组与C组的FL、FW、SGR、WGR和FCR均无显著差异(P>0.05)。L组的FL显著高于C组(P<0.05), 但其他指标无显著差异(P>0.05)。M组FL、FW、SGR和WGR均显著高于C组(P<0.05)。在所有流速处理组中, M组表现出最佳的生长性能, 其FL、FW、SGR和WGR均显著高于H组, 但与L组无显著差异(P>0.05)。与其他指标相反, FCR呈现先下降后上升的趋势, H流速组的FCR显著高于L流速组(P<0.05), 但与C组及M组相比没有显著差异(P>0.05)。
表 1 不同流速对许氏平鲉生长的影响Table 1. The effect of different flow velocities on growth of Sebastes schlegelii生长指标Growth index 流速Flow velocity C (0) L (0.5 BL/s) M (1.5 BL/s) H (2.5 BL/s) 终体长FL (cm) 12.58±0.11a 13.12±0.10b 13.42±0.23b 12.46±0.05a 终体重FW (g) 54.32±0.99a 58.82±1.16ab 62.04±2.10 b 54.43±1.29a 特定生长率SGR (%/d) 2.13±0.03a 2.31±0.10ab 2.45±0.04b 2.13±0.04a 增重率WGR (%) 0.99±0.13a 1.19±0.12ab 1.37±0.09b 1.00±0.04a 饲料系数FCR 2.83±0.20ab 2.22±0.26a 2.4±0.25ab 3.05±0.08b 注: 均值后不同上标字母表示同一时间段不同流速组间具有显著性差异(P<0.05)Note: Different superscripts after means indicate significant differences (P<0.05) 2.2 流速对许氏平鲉行为的影响
本研究测定了8个行为参数, 按照行为表现的特征, 将其归纳为3类行为状态指标, 分别为个体的运动参数、偏好区域和活动状态。
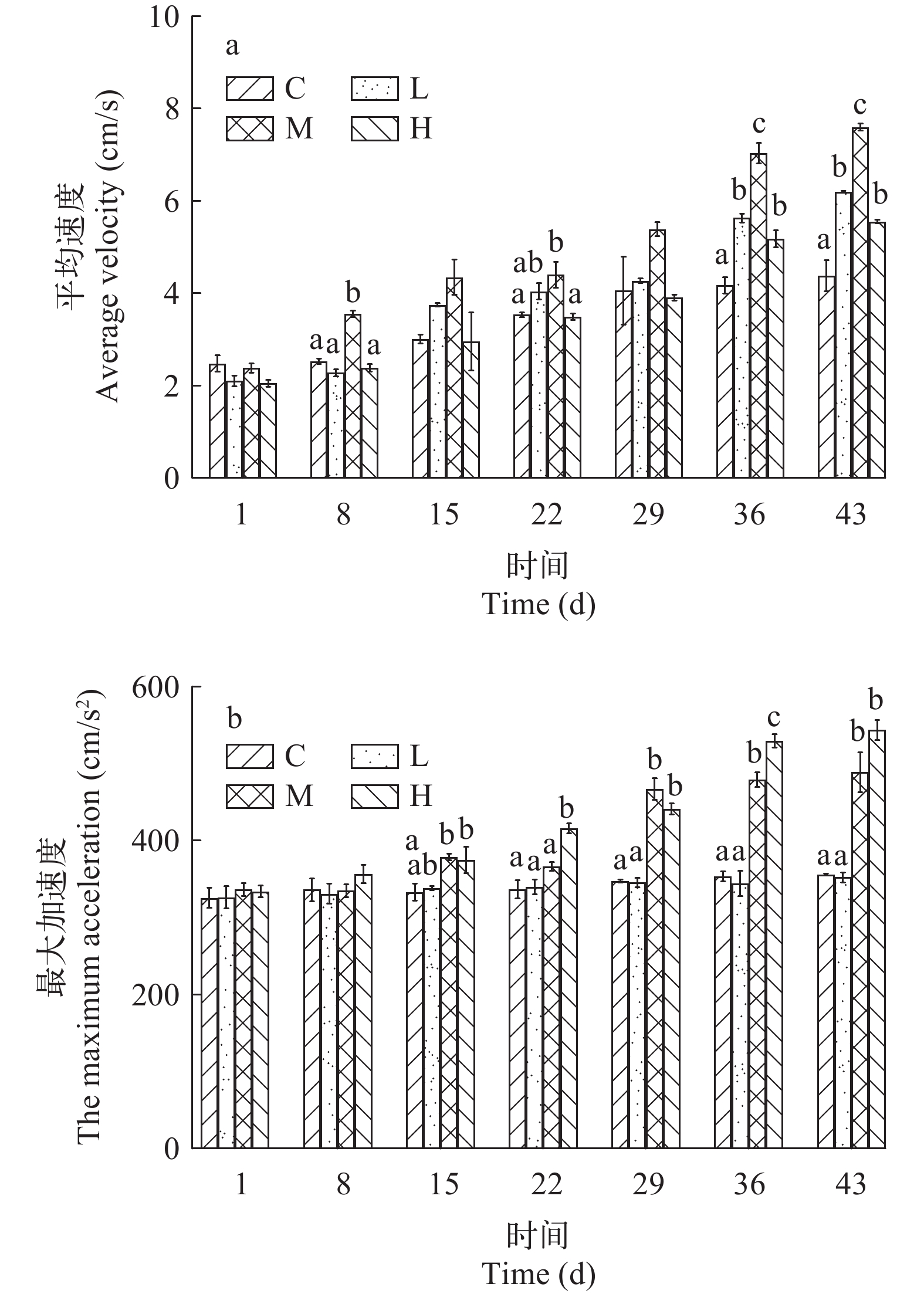
流速对许氏平鲉运动参数的影响 如图 3a所示, 随实验时间增加, 许氏平鲉的平均速度均呈现上升的趋势, 但各流速组的上升趋势不同。C组在整个养殖期间内的平均速度相对平稳。对于3个流速处理组来说, 许氏平鲉的平均速度均随时间增加呈现显著上升的趋势。在实验后期, M组许氏平鲉的平均速度显著高于L组和H组(P<0.05), L组与H组之间无显著差异(P>0.05), C组显著低于3个流速处理组(P<0.05)。
![]() 图 3 不同流速对许氏平鲉运动参数的影响误差线上方不同字母表示差异显著(P<0.05); 下同Figure 3. Variation in movement parameters of Sebastes schlegelii at different flow velocitiesDifferent letters above the error bars indicate significant differences (P<0.05); the same applies below
图 3 不同流速对许氏平鲉运动参数的影响误差线上方不同字母表示差异显著(P<0.05); 下同Figure 3. Variation in movement parameters of Sebastes schlegelii at different flow velocitiesDifferent letters above the error bars indicate significant differences (P<0.05); the same applies below如图 3b所示, C组和L组许氏平鲉的最大加速度在整个实验期间没有显著变化(P>0.05)。M组和H组的最大加速度随实验时间的延长均呈现显著上升趋势。在实验30d后, M组和H组幼鱼的最大加速度显著高于C组和L组(P<0.05), 但M组和H组, C组与L组之间无显著差异(P>0.05)。
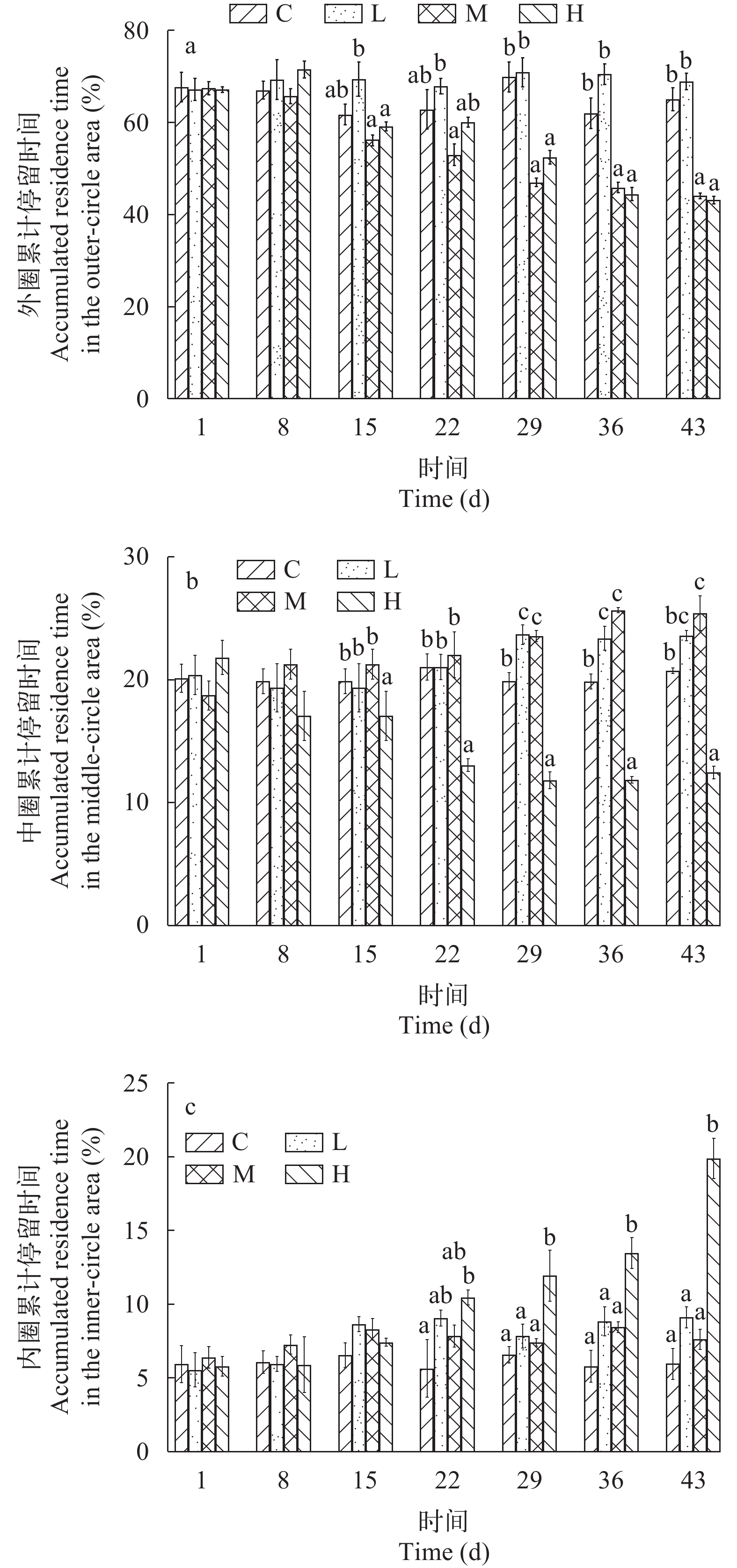
流速对许氏平鲉偏好区域的影响 如图 4a所示, C组和L组许氏平鲉的外圈累计停留时间在整个实验期间没有显著差异(P>0.05)。M组和H组幼鱼在外圈的累计停留时间随时间增加呈现显著下降的趋势。在实验30d后, M组和H组许氏平鲉的外圈累计停留时间显著低于C组和L组(P<0.05), 但M组与H组之间无显著差异(P>0.05), C组与L组之间无显著差异(P>0.05)。
![]() 图 4 不同流速对许氏平鲉偏好区域的影响Figure 4. Variation in preferred areas of Sebastes schlegelii at different flow velocities
图 4 不同流速对许氏平鲉偏好区域的影响Figure 4. Variation in preferred areas of Sebastes schlegelii at different flow velocities如图 4b所示, C组和L组许氏平鲉的中圈累计停留时间在整个实验期间没有显著差异(P>0.05)。M组的中圈累计停留时间随时间增加平稳上升, 而H组随时间增加呈现显著下降的趋势。其中, 自实验15d起, H组许氏平鲉的中圈累计停留时间显著低于其他组(P<0.05); 在实验30d后, C组的中圈累计停留时间显著低于M组(P<0.05), L组与M组无显著差异(P>0.05)。
如图 4c所示, C组和M组许氏平鲉的内圈累计停留时间在整个实验期间没有显著差异(P>0.05)。L组的内圈累计停留时间随时间增加缓慢上升, 而H组随时间增加呈显著上升的趋势。在实验30d后, H组许氏平鲉的内圈累计停留时间显著高于其他处理组(P<0.05), 且这3组之间没有显著差异(P>0.05)。
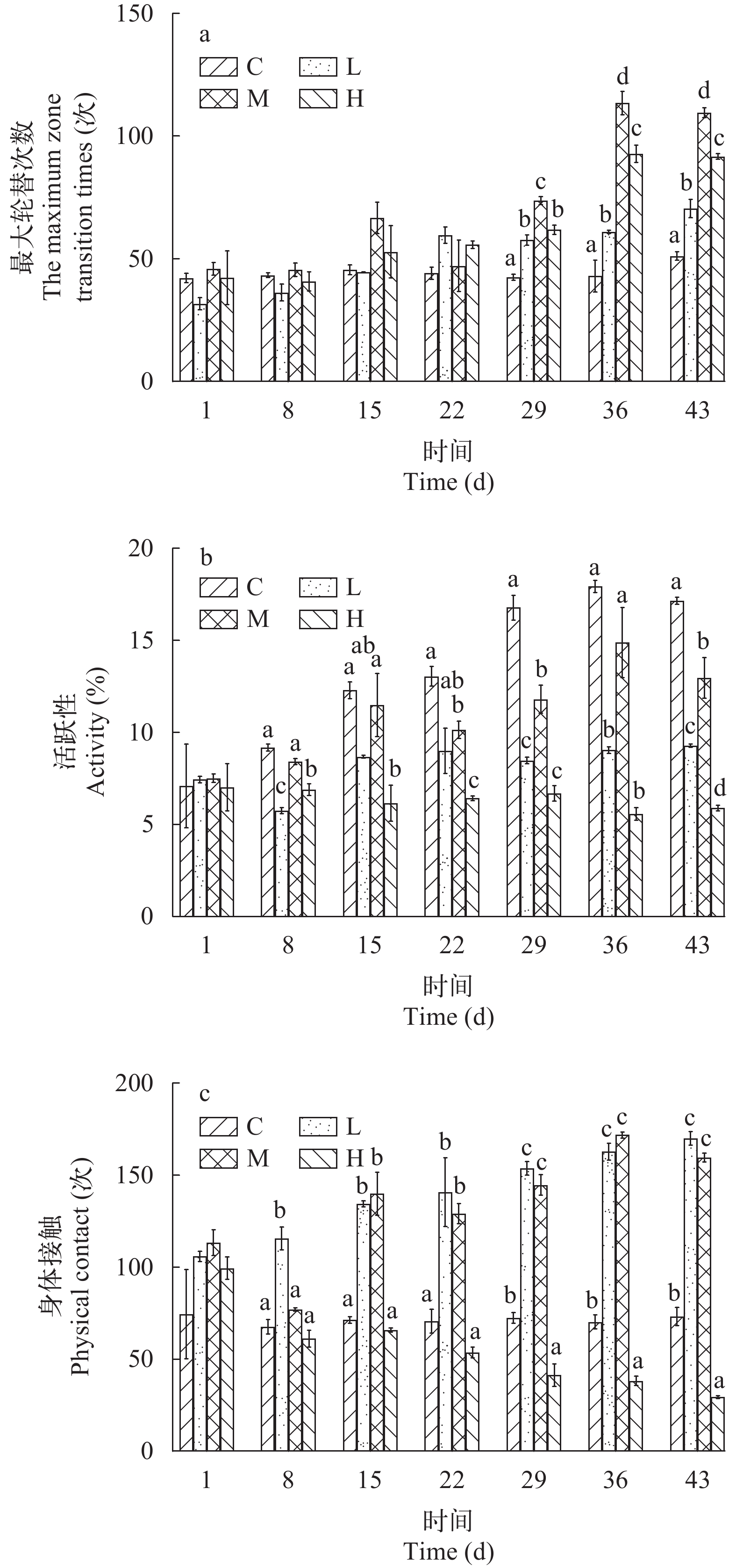
流速对许氏平鲉活动状态的影响 如图 5a所示, C组许氏平鲉的最大轮替次数在整个实验期间没有显著差异(P>0.05)。对于3个流速处理组来说, 许氏平鲉的最大轮替次数均随时间增加呈现显著上升的趋势。在实验30d后, C组许氏平鲉的最大轮替次数显著低于其他有流速的处理组(P<0.05), M组显著高于其他3组(P<0.05), 并且, H组的最大轮替次数显著高于L组(P<0.05)。
![]() 图 5 不同流速对许氏平鲉活动状态的影响Figure 5. Variation in activity status of Sebastes schlegelii at different flow velocities
图 5 不同流速对许氏平鲉活动状态的影响Figure 5. Variation in activity status of Sebastes schlegelii at different flow velocities如图 5b所示, H组许氏平鲉的活跃性在整个实验期间没有显著差异(P>0.05)。C组的活跃性随时间显著上升, M组的活跃性随时间增加呈现上升趋势, L组呈现缓慢上升趋势。自实验30d开始, L组和H组许氏平鲉的活跃性始终显著低于C组和M组(P<0.05); 在实验结束时, C组的活跃性显著高于其他有流速处理组(P<0.05)。
如图 5c所示, C组的身体接触在整个实验期间没有显著差异(P>0.05)。对于3个流速组来说, L组和M组许氏平鲉的身体接触随时间增加呈显著上升的趋势, H组的身体接触随时间增加呈显著下降的趋势。其中, 在实验15d后, L组和M组许氏平鲉的身体接触显著高于其他处理组(P<0.05), 且L组与M组之间无显著差异(P>0.05); 在实验30d后, C组的身体接触显著高于H组(P<0.05), H组显著低于其他处理组(P<0.05)。
3. 讨论
3.1 流速对许氏平鲉生长的影响
优化养殖环境可以为养殖鱼类的性能和福利带来明显的益处, 提高水产养殖的整体效益[24]。以往的研究结果表明, 适宜的流速环境能够在促进鱼类健康和福利情况的同时, 提高鱼类的生产性能[25, 26]。本研究结果显示, L组和M组许氏平鲉的生长性能显著提高, 其中M组的FL、FW、SGR和WGR显著高于其他处理组, 表现出最佳的生长性能, 这说明当流速小于1.5 BL/s时, 随着流速的增加, 许氏平鲉的生长出现上升趋势。相似的结果也在大鳞大麻哈鱼(Oncorhynchus tshawytscha) [3]、虹鳟(Oncorhynchus mykiss) [27]和大西洋鲑(Salmo salar) [28]等的研究中被证明。这一方面可能是由于适宜的水流诱导鱼进行游泳运动, 减少了在低流速条件下鱼类由于频繁移动和竞争行为所消耗的能量[13, 29—31]; 另一方面, 适宜的水流速度为缓解鱼类压力、提高摄食欲望和抗病能力提供了帮助, 并最终表现为较高的免疫能力与生长性能[31, 32]。
H组许氏平鲉的各项生长指标出现下降, 这可能是因为2.5 BL/s接近许氏平鲉的最大可持续游动速度, 而为了适应高流速, 许氏平鲉不得不保持较高的游泳速度, 这使得原本将用于生长的能量转移到了运动上[31, 33]; 另一方面, 过高流速刺激鱼类被迫进行高强度的运动, 从而引起鱼体的应激反应和游泳疲劳, 抑制了许氏平鲉的生长[14, 34—36]。史航等[37]的研究表明, 体长为10—22 cm的许氏平鲉相对临界游速为3.11 BL/s, 在本研究中, 规格较小的许氏平鲉幼鱼的最佳生长性能出现在约1.5 BL/s的流速条件下, 也符合这一趋势, 这可能与许氏平鲉为近海底层鱼类, 其喜爱在岩礁附近生活的习性有关[38, 39]。本研究结果显示, 在1.0—1.5 BL/s流速下, 许氏平鲉能够获得最佳的生长性能。
3.2 流速对许氏平鲉行为的影响
研究结果显示, M组的运动参数(平均速度和最大加速度)显著高于其他处理组。Robinson等[40]的研究表明, 在转移入养殖网箱前系统地对养殖鱼类进行游泳训练, 可以提高它们的游泳能力。在实验期间, 1.5 BL/s的水流训练提高了流速组许氏平鲉的游泳能力, 且降低了不利于其生长的自发性行为; 相对的, C组许氏平鲉活跃性在所有处理中最高, 而其在各圈层之间的最大轮替次数却最少, 证明了在静水条件下, 许氏平鲉会进行着短距离、高频率的自发活动并损失能量。除此之外, Hvas等[41]的研究结果表明, 适宜的流速能够提高鱼群灵活变换位置及调整群体结构的能力, 以应对短暂的水流游泳挑战, 这是一种良好养殖福利的象征。从这一点上看, 流速刺激生长而带来的一部分益处可能是由于流速诱导的行为效应而产生的。Johansson等[42]的研究也表明, 当大西洋鲑面对水流时, 圆形群体游泳结构发生了短暂的中断, 表明鱼类具有高度的灵活性, 可以充分适应海水网箱环境的突然变化。结果显示, 处于M组流速下的许氏平鲉, 其活动状态(最大轮替次数、活跃性和身体接触)显著高于其他处理组, 这可能是因为1.5 BL/s与许氏平鲉的最适游速相接近, 适宜的流速帮助许氏平鲉更好地应对压力挑战[43], 促进了许氏平鲉形成有序的群体结构, 并能够及时调整来适应流速环境以及保持群体稳定性[13, 41]。
生活在流水中的鱼类大多具有趋流性。它们能根据水流的速度随时调整自身的游速, 使自身保持逆流游泳状态或长时间地停留在某一特定的位置[42, 44]。Hvas等[44]研究表明, 养殖的大西洋鲑鱼在低流速下会以自主选择的游泳速度在网箱中绕圈游泳以避免相互碰撞以及与网箱壁碰撞; 而当环境水流速度超过了这个自发的游泳速度, 鲑将停止巡航游泳, 转变为与环境速度适应的游泳速度来保持在水流中的位置。结果显示, M组许氏平鲉幼鱼主要分布于养殖区域中圈, 这可能是许氏平鲉主动选择了其最适的流速偏好范围, 而该范围与中圈和内圈、外圈的区域交接处相重叠, 许氏平鲉始终在此徘徊游动, 所以M组区域间的最大轮替次数显著高于其他处理。而H组的区域间的最大轮替次数较高, 可能是因为H组许氏平鲉在实验开始后面对高流速开始由外圈及中圈向内圈转移, 并最终停留于内圈, 也说明了许氏平鲉在高流速条件下, 努力通过改变其所在的区域位置, 寻求其喜爱的偏好流速区域来应对过高流速的冲击[42, 45]。Georgopoulou等[46]关于鱼类对水箱表面的偏好研究与我们的研究结果相似, 表明区域偏好可能是表征许氏平鲉行为表现的特征指标。
如果水流速度继续增大, 鱼的游泳状态将从稳定游泳变为更依赖爆发和滑动游泳[45]。研究结果显示, 在实验后期, H组的最大加速度最高, 说明2.5 BL/s的水流速度超过了许氏平鲉游泳能力, 其主要依赖爆发游泳以保持在水中的相对位置[41]。Hvas等[36]研究表明, 网箱内的水流速度一旦超过鱼类的自愿游泳速度, 将会损害鱼类表达其自然行为的能力。H组许氏平鲉的活跃性和身体接触显著低于其他处理组。这可能是因为处于H组流速下的许氏平鲉为了适应高流速的水流, 被迫提高了游泳速度, 形成抵抗水流的群体模式[45], 导致许氏平鲉正常的行为模式被抑制, 所以呈现出较低的活跃性和身体接触[14]。活跃性、身体接触和区域间轮替次数等活动状态显著受到流速变化的影响。因此, 以上特征也可以用作评价许氏平鲉应对水流的行为响应指标。
综上所述, 在集约化养殖条件下, 许氏平鲉养殖池的水流速度设定在1.0—1.5 BL/s, 能够促使其获得最佳生长性能, 并保障其表现出更自然的行为能力。我们可以通过观测许氏平鲉的区域偏好指标以及包括活跃性、身体接触和区域间轮替次数在内的活动状态指标, 作为其所处流速环境状态的行为特征的判断依据, 并以此来推进许氏平鲉养殖过程中行为的识别, 促进养殖过程管理的精细化和智能化程度。
-
![]()
图 2 采样点及分析区示意图
a. 养殖桶侧视图; b. 养殖桶俯视图
Figure 2. Schematic diagram of measurement points and analysis areas
a. lateral view of rearing tank; b. top view of rearing tank
![]()
图 3 不同流速对许氏平鲉运动参数的影响
误差线上方不同字母表示差异显著(P<0.05); 下同
Figure 3. Variation in movement parameters of Sebastes schlegelii at different flow velocities
Different letters above the error bars indicate significant differences (P<0.05); the same applies below
![]()
图 4 不同流速对许氏平鲉偏好区域的影响
Figure 4. Variation in preferred areas of Sebastes schlegelii at different flow velocities
![]()
图 5 不同流速对许氏平鲉活动状态的影响
Figure 5. Variation in activity status of Sebastes schlegelii at different flow velocities
表 1 不同流速对许氏平鲉生长的影响
Table 1 The effect of different flow velocities on growth of Sebastes schlegelii
生长指标Growth index 流速Flow velocity C (0) L (0.5 BL/s) M (1.5 BL/s) H (2.5 BL/s) 终体长FL (cm) 12.58±0.11a 13.12±0.10b 13.42±0.23b 12.46±0.05a 终体重FW (g) 54.32±0.99a 58.82±1.16ab 62.04±2.10 b 54.43±1.29a 特定生长率SGR (%/d) 2.13±0.03a 2.31±0.10ab 2.45±0.04b 2.13±0.04a 增重率WGR (%) 0.99±0.13a 1.19±0.12ab 1.37±0.09b 1.00±0.04a 饲料系数FCR 2.83±0.20ab 2.22±0.26a 2.4±0.25ab 3.05±0.08b 注: 均值后不同上标字母表示同一时间段不同流速组间具有显著性差异(P<0.05)Note: Different superscripts after means indicate significant differences (P<0.05)  下载: 导出CSV
下载: 导出CSV
-
[1] 任效忠, 王江竹, 薛博茹, 等. 方形圆弧角海水养殖池排污特性的试验研究 [J]. 海洋环境科学, 2021, 40(5): 790-797. doi: 10.12111/j.mes.20200166 Ren X Z, Wang J Z, Xue B R, et al. Experimental study on the blowdown characteristics of square arc-angle maricultural tank [J]. Marine Environmental Science, 2021, 40(5): 790-797. doi: 10.12111/j.mes.20200166
[2] 车宗龙, 任效忠, 张倩. 循环水养殖系统中水动力特性及其与鱼类相互影响研究进展 [J]. 大连海洋大学学报, 2021, 36(5): 886-898. doi: 10.16535/j.cnki.dlhyxb.2021-104 Che Z L, Ren X Z, Zhang Q. Research progress on hydrodynamics in a recirculating aquaculture tank system: a review [J]. Journal of Dalian Ocean University, 2021, 36(5): 886-898. doi: 10.16535/j.cnki.dlhyxb.2021-104
[3] Parker T M, Barnes M E. Rearing velocity impacts on landlocked fall Chinook salmon (Oncorhynchus tshawytscha) growth, condition, and survival [J]. Open Journal of Animal Sciences, 2014, 4(5): 244-252. doi: 10.4236/ojas.2014.45031
[4] Nilsen A, Hagen Ø, Johnsen C A, et al. The importance of exercise: Increased water velocity improves growth of Atlantic salmon in closed cages [J]. Aquaculture, 2019(501): 537-546.
[5] Waldrop T, Summerfelt S, Mazik P, et al. The effects of swimming exercise and dissolved oxygen on growth performance, fin condition and precocious maturation of early-rearing Atlantic salmon Salmo salar [J]. Aquaculture Research, 2018, 49(2): 801-808. doi: 10.1111/are.13511
[6] Arbeláez-Rojas G A, Moraes G, da Silva Nunes C, et al. Sustained swimming mitigates stress in juvenile Brycon amazonicus reared in high stocking densities [J]. Pesquisa Agropecuária Brasileira, 2017, 52(1): 1-9.
[7] 刘梅, 练青平, 倪蒙, 等. 池塘内循环流水养殖模式对大口黑鲈生长性能、抗氧化酶、消化酶及消化道组织结构和菌群的影响 [J]. 水产学报, 2021, 45(12): 2011-2028. Liu M, Lian Q P, Ni M, et al. Effects of inner-pond raceway aquaculture on the growth performance, antioxidant enzymes, digestive enzymes, digestive tract structure, and bacterial flora of largemouth bass (Micropterus salmoides) [J]. Journal of Fisheries of China, 2021, 45(12): 2011-2028.
[8] 虞顺年, 魏小岚, 韦芳三, 等. 不同运动强度对黑鲷生长、血清和肝脏抗氧化指标的影响 [J]. 水生生物学报, 2018, 42(02): 255-263. doi: 10.7541/2018.032 Yu S N, Wei X L, Wei F S, et al. Effects of different exercise intensity on growth and serum and liver antioxidant indices of Sparus macrocephalus [J]. Acta Hydrobiologica Sinica, 2018, 42(02): 255-263. doi: 10.7541/2018.032
[9] Li X, Ji L, Wu L, et al. Effect of flow velocity on the growth, stress and immune responses of turbot (Scophthalmus maximus) in recirculating aquaculture systems [J]. Fish & Shellfish Immunology, 2019(86): 1169-1176.
[10] Liu G, Wu Y, Qin X, et al. The effect of aerobic exercise training on growth performance, innate immune response and disease resistance in juvenile Schizothorax prenanti [J]. Aquaculture, 2018(486): 18-25.
[11] Chen Z, Ye Z, Ji M, et al. Effects of flow velocity on growth and physiology of juvenile largemouth bass (Micropterus salmoides) in recirculating aquaculture systems [J]. Aquaculture Research, 2021, 52(7): 3093-3100. doi: 10.1111/are.15153
[12] Montgomery J, Carton G, Voigt R, et al. Sensory processing of water currents by fishes [J]. Philosophical Transactions of the Royal Society of London Series B: Biological Sciences, 2000, 355(1401): 1325-1327. doi: 10.1098/rstb.2000.0693
[13] Solstorm F, Solstorm D, Oppedal F, et al. Not too slow, not too fast: water currents affect group structure, aggression and welfare in post-smolt Atlantic salmon Salmo salar [J]. Aquaculture Environment Interactions, 2016(8): 339-347.
[14] Hvas M, Folkedal O, Oppedal F. What is the limit of sustained swimming in Atlantic salmon post smolts [J]? Aquaculture Environment Interactions, 2021(13): 189-198.
[15] Weirup L, Schulz C, Seibel H, et al. Scale cortisol is positively correlated to fin injuries in rainbow trout (Oncorhynchus mykiss) reared in commercial flow through systems [J]. Aquaculture, 2021(543): 736924.
[16] Solstorm F, Solstorm D, Oppedal F, et al. Fast water currents reduce production performance of post-smolt Atlantic salmon Salmo salar [J]. Aquaculture Environment Interactions, 2015, 7(2): 125-134. doi: 10.3354/aei00143
[17] Macaulay G, Bui S, Oppedal F, et al. Challenges and benefits of applying fish behaviour to improve production and welfare in industrial aquaculture [J]. Reviews in Aquaculture, 2021, 13(2): 934-948. doi: 10.1111/raq.12505
[18] An D, Huang J, Wei Y. A survey of fish behaviour quantification indexes and methods in aquaculture [J]. Reviews in Aquaculture, 2021, 13(4): 2169-2189. doi: 10.1111/raq.12564
[19] Waldrop T, Summerfelt S, Mazik P, et al. The effects of swimming exercise and dissolved oxygen on growth performance, fin condition and survival of rainbow trout Oncorhynchus mykiss [J]. Aquaculture Research, 2020, 51(6): 2582-2589. doi: 10.1111/are.14600
[20] Xi D, Zhang X, Lü H, et al. Cannibalism in juvenile black rockfish, Sebastes schlegelii (Hilgendorf, 1880), reared under controlled conditions [J]. Aquaculture, 2017(479): 682-689.
[21] Reiser S, Sähn N, Pohlmann D M, et al. Rearing juvenile brown Salmo trutta (L.), and rainbow trout Oncorhynchus mykiss (Walbaum), in earthen ponds with and without an induced current [J]. Journal of Applied Aquaculture, 2021, 33(1): 32-52. doi: 10.1080/10454438.2019.1678545
[22] Martling S R, Voorhees J M, Huysman N, et al. Juvenile Fall Chinook Salmon Rearing Performance is Not Affected by Exercise and Structure [C]. Proceedings of the Proceedings of the South Dakota Academy of Science. South Dakota. 2021: 100.
[23] Silva A T, Bærum K M, Hedger R D, et al. The effects of hydrodynamics on the three-dimensional downstream migratory movement of Atlantic salmon [J]. Science of the Total Environment, 2020(705): 135773.
[24] Arechavala-Lopez P, Cabrera-Álvarez M J, Maia C M, et al. Environmental enrichment in fish aquaculture: a review of fundamental and practical aspects [J]. Reviews in Aquaculture, 2022, 14(2): 704-728. doi: 10.1111/raq.12620
[25] Inoue L A K A, Hackbarth A, Arberláez-Rojas G, et al. Growth performance and metabolism of the Neotropical fish Piaractus mesopotamicus under sustained swimming [J]. Aquaculture, 2019(511): 734219.
[26] Shrivastava J, Rašković B, Blust R, et al. Exercise improves growth, alters physiological performance and gene expression in common carp (Cyprinus carpio) [J]. Comparative Biochemistry and Physiology Part A:Molecular & Integrative Physiology, 2018(226): 38-48.
[27] Voorhees J M, Huysman N, Krebs E, et al. Use of exercise and structure during rainbow trout rearing [J]. Open Journal of Applied Sciences, 2020, 10(5): 258-269. doi: 10.4236/ojapps.2020.105020
[28] Timmerhaus G, Lazado C C, Cabillon N A R, et al. The optimum velocity for Atlantic salmon post-smolts in RAS is a compromise between muscle growth and fish welfare [J]. Aquaculture, 2021(532): 736076.
[29] Fernö A, Bjelland O, Kristiansen T S. The Diverse World of Fishes [M]. Animal Welfare. Cham: Springer International Publishing, 2020: 43-72.
[30] Balseiro P, Moe Ø, Gamlem I, et al. Comparison between Atlantic salmon Salmo salar post-smolts reared in open sea cages and in the Preline raceway semi-closed containment aquaculture system [J]. Journal of Fish Biology, 2018, 93(3): 567-579. doi: 10.1111/jfb.13659
[31] Ashley P J. Fish welfare: current issues in aquaculture [J]. Applied Animal Behaviour Science, 2007, 104(3/4): 199-235.
[32] Zhu Z M, Song B L, Lin X T, et al. Effects of water-current speed on hematological, biochemical and immune parameters in juvenile tinfoil barb, Barbonymus schwanenfeldii (Bleeker, 1854) [J]. Chinese Journal of Oceanology and Limnology, 2016, 34(1): 118-124. doi: 10.1007/s00343-015-4276-6
[33] Morro B, Davidson K, Adams T P, et al. Offshore aquaculture of finfish: big expectations at sea [J]. Reviews in Aquaculture, 2022, 14(2): 791-815. doi: 10.1111/raq.12625
[34] Nilsen A, Nielsen K V, Bergheim A. A closer look at closed cages: growth and mortality rates during production of post-smolt Atlantic salmon in marine closed confinement systems [J]. Aquacultural Engineering, 2020(91): 102124.
[35] Is-Haak J, Kaewnern M, Yoonpundh R, et al. Oxygen consumption rates of hybrid red tilapia at different sizes during challenge to water velocity [J]. Journal of Fisheries, 2019, 43(2): 52-65.
[36] Hvas M, Oppedal F. Sustained swimming capacity of Atlantic salmon [J]. Aquaculture Environment Interactions, 2017(9): 361-369.
[37] 史航, 陈勇, 赵子仪, 等. 许氏平鲉、大泷六线鱼临界游速与爆发游速及其生理指标的研究 [J]. 大连海洋大学学报, 2010, 25(5): 407-412. doi: 10.3969/j.issn.1000-9957.2010.05.006 Shi H, Chen Y, Zhao Z Y, et al. Critical swimming speed, burst swimming speed and related physiological index in rockfish Sebastes schlegeli and fat greenling Hexagrammos otakii [J]. Journal of Dalian Ocean University, 2010, 25(5): 407-412. doi: 10.3969/j.issn.1000-9957.2010.05.006
[38] 席杨, 田涛, 杨军, 等. 许氏平鲉幼鱼在不同流场环境下的趋礁行为研究 [J]. 大连海洋大学学报, 2020, 35(3): 399-406. doi: 10.16535/j.cnki.dlhyxb.2019-313 Xi Y, Tian T, Yang J, et al. Reef approaching behavior of juvenile Sebastes schlegeli under different flow fields [J]. Journal of Dalian Ocean University, 2020, 35(3): 399-406. doi: 10.16535/j.cnki.dlhyxb.2019-313
[39] 刘鸿雁, 吕洪斌, 张沛东, 等. 人工鱼礁模型和大型海藻对许氏平鲉和大泷六线鱼幼鱼的诱集作用 [J]. 水产学报, 2018, 42(1): 48-59. Liu H Y, Lü H B, Zhang P D, et al. Attraction effect of artificial reef model and macroalgae on juvenile Sebastes schlegelii and Hexagrammos otakii [J]. Journal of Fisheries of China, 2018, 42(1): 48-59.
[40] Robinson N A, Timmerhaus G, Baranski M, et al. Training the salmon’s genes: influence of aerobic exercise, swimming performance and selection on gene expression in Atlantic salmon [J]. BMC Genomics, 2017, 18(1): 971. doi: 10.1186/s12864-017-4361-7
[41] Hvas M, Folkedal O, Oppedal F. Fish welfare in offshore salmon aquaculture [J]. Reviews in Aquaculture, 2021, 13(2): 836-852. doi: 10.1111/raq.12501
[42] Johansson D, Laursen F, Fernö A, et al. The interaction between water currents and salmon swimming behaviour in sea cages [J]. PLoS One, 2014, 9(5): e97635. doi: 10.1371/journal.pone.0097635
[43] Roza e Silva M L R E, Pereira R T, Arvigo A L, et al. Effects of water flow on ventilation rate and plasma cortisol in Nile tilapia introduced into novel environment [J]. Aquaculture Reports, 2020(18): 100531. doi: 10.1016/j.aqrep.2020.100531
[44] Hvas M, Folkedal O, Solstorm D, et al. Assessing swimming capacity and schooling behaviour in farmed Atlantic salmon Salmo salar with experimental push-cages [J]. Aquaculture, 2017(473): 423-429. doi: 10.1016/j.aquaculture.2017.03.013
[45] Johannesen Á, Patursson Ø, Kristmundsson J, et al. Waves and currents decrease the available space in a salmon cage [J]. PLoS One, 2022, 17(2): e0263850. doi: 10.1371/journal.pone.0263850
[46] Georgopoulou D G, Stavrakidis-Zachou O, Mitrizakis N, et al. Tracking and analysis of the movement behavior of European seabass (Dicentrarchus labrax) in aquaculture systems [J]. Frontiers in Animal Science, 2021(2): 754520. doi: 10.3389/fanim.2021.754520
-
期刊类型引用(4)
1. 房鼎航,魏涛,贾彪,韩岩珂,田涛,于晓明. 流速对许氏平鲉幼鱼生长、个性和焦虑行为的影响. 安徽农学通报. 2025(01): 52-58 .  百度学术
百度学术
2. 张佳,葛晓雨,陈松猛,张旭,李海霞,王婕,马真. 投喂频率对红鳍东方鲀生长和行为的影响. 水生生物学报. 2024(01): 77-85 . 本站查看
3. 关兴宇,张雨田,邓立宇,吴忠鑫,于晓明,田涛. 流速对许氏平鲉群体行为和生理机能的影响. 安徽农学通报. 2024(10): 40-46 . 百度学术
4. 张雨田,田涛,于晓明,吴忠鑫,李青霞,高东奎. 光照颜色对许氏平鲉幼鱼行为和生理的影响. 上海海洋大学学报. 2024(06): 1392-1405 . 百度学术
其他类型引用(4)
计量
- 文章访问数: 1125
- HTML全文浏览量: 537
- PDF下载量: 55
- 被引次数: 8