
THE OCCURRENCE OF AHPND IN POND CULTURED LITOPENAEUS VANNAMEI AND ITS INFLUENCE ON ENVIRONMENT, PATHOGEN AND IMMUNE FACTORS
-
摘要: 为研究虾急性肝胰腺坏死病(Acute Hepatopancreas Necrosis Disease, AHPND)的发生与环境、病原和虾体免疫间的相互关系, 文章对池塘养殖凡纳滨对虾(Litopenaeus vannamei)AHPND发生及其环境、病原、虾体免疫因子进行持续性跟踪监测。结果表明, 试验点的气温、水温、溶解氧(DO)、pH、盐度、氨氮(NH4-N)和亚硝态氮(NO2-N)波动范围为21—29℃、24.8—31℃、1.4—8.32 mg/L、8—8.91、34—50、0.01—0.26 mg/L和0.005—0.212 mg/L; 水体可培养细菌和弧菌数量变化范围为3×103—2.4×105和2×102—1.8×104 CFU/mL, 虾体肝胰腺内可培养细菌和弧菌数量变化范围为9.8×104—8.8×106和3.9×103—3.61×106 CFU/g; 16S rDNA鉴定结果显示, 在可培养优势菌株中, 弧菌检出数量达到135株, 占全部鉴定菌株的61.6%, 其中主要弧菌种类有欧文氏弧菌(Vibrio owensii)、坎贝氏弧菌(V. campbellii)、副溶血弧菌(V. parahaemolyticus)、溶藻弧菌(V. alginolyticus)和哈维氏弧菌(V. harveyi); 虾体中ACP、AKP、SOD、LZM和PO等免疫酶活的变化范围分别为7.5—75、1—8.5、2.4—11.07、1.3—43和6.23—28 U/mg。结合AHPND发生前后各理化因子变化的相关性分析表明, 水温、虾体肝胰腺内可培养细菌和弧菌数量、DO、LZM和PO可作为池塘养殖模式下对虾警示AHPND的指示因子。相关研究结果为指导池塘养殖凡纳滨对虾健康养殖及AHPND临床防控提供数据支撑和科学依据。Abstract: Acute Hepatopancreatic Necrosis Disease (AHPND) is an important limiting factor affecting the healthy cultivation of shrimp in recent years, which has brought huge economic losses to the global shrimp industry. In order to explore the relationship between the occurrence of AHPND and environment, pathogen and shrimp immunity factors, the pond cultured Litopenaeus vannamei systems were continuously monitored. Based on the 35 days continuously monitoring showed that, the air temperature, water temperature, dissolved oxygen (DO), pH, salinity, ammonia nitrogen (NH4-N) and nitrite (NO2-N) were fluctuated in the range of 21—29℃, 24.8—31℃, 1.4—8.32 mg/L, 8—8.91, 34—50, 0.01—0.26 mg/L and 0.005—0.212 mg/L, respectively; the culturable bacteria and Vibrio sp. in water and shrimp hepatopancreas were fluctuated in the range of 3×103—2.4×105 CFU/mL, 2×102—1.8×104 CFU/mL, 9.8×104—8.8×106 and 3.9×103—3.61×106 CFU/g, respectively; 135 isolates of Vibrio sp. were detected based on 16S rDNA, and the main Vibrio species was V. owensii, V. campbellii, V. parahaemolyticus, V. alginolyticus and V. harveyi; the activities of ACP, AKP, SOD, LZM and PO were 7.5—75, 1.0—8.5, 2.4—11.07, 1.3—43 and 23—28 U/mg, respectively. Combined the physical and chemical factors before and after the occurrence of AHPND, the results showed that, water temperature, culturable bacteria and Vibrio sp. in hepatopancreas, DO, LZM and PO could be used as the early warning candidate factors for the occurrence of AHPND in pond culture. The results provide a scientific basis for the healthy culture of L. vannamei and the prevention and control of AHPND disease.
-
Keywords:
- AHPND /
- Environmental factors /
- Culturable bacteria /
- Immune factors /
- Early warning factors /
- Litopenaeus vannamei
-
凡纳滨对虾(Litopenaeus vannamei)生长速度快、耐高温、抗病力强, 是我国对虾养殖的优良品种, 同时也是世界养殖产量最高的三种优良对虾种类之一。但对虾养殖业的快速发展和不规范运作导致对虾的病害问题日趋严重, 为对虾养殖产业带来巨大的经济损失。对虾病害具有暴发性强、死亡率高和控制困难等特点。且对虾养殖过程中病害发生区域广、病害问题复杂、病原种类繁多, 严重制约着对虾养殖业的健康稳定发展[1-3]。近几年来, 对虾急性肝胰腺坏死病(Acute Hepatopancreas Necrosis Disease, AHPND)造成大批对虾发病死亡, 给全球对虾养殖业带来巨大经济损失[4, 5]。迄今为止, 已报道的AHPND致病原复杂多样, 包括副溶血弧菌(Vibrio parahemolyticus)、哈维氏弧菌(V. harveyi)、欧文氏弧菌(V. owensii)、坎贝氏弧菌(V. campbellii)和溶藻弧菌(V. alginolyticus)等[6-10]。同时, 针对AHPND的临床和病理特征、疾病发生机制、分子检测技术等均开展了大量的基础理论研究且获得阶段性研究成果[11-13]。
病害发生是由环境因子、病原微生物和机体自身免疫力三要素相互作用的结果, 其中病原是引发水产动物疫病发生的最主要因素, 也是病害预防研究的关键内容。养殖环境变化是多种疾病发生的诱因, 环境因子的变化既影响病原体生存代谢及其对宿主的侵袭能力, 同时也影响宿主的免疫力, 是病害发生发展的重要调控因素[14, 15]。对虾病害发生与其生存的生态环境(EV/CV, Environmental and climatic variability)息息相关, 水体中的pH、DO、温度和盐度等环境因子对其生长、发育和繁殖都有着重要的调控作用。目前, 对于池塘养殖对虾的疾病发生与环境因子的相互关系还缺乏全面深入的研究。通过多种因子的联合分析, 全面了解池塘养殖模式下凡纳滨对虾养殖系统内的环境因子、微生物因子和健康指示因子的级联关系, 掌握疾病发生与各因素的相互关系, 对控制和预防池塘养殖凡纳滨对虾疾病的发生具有重要的指导意义。
本研究以河北省沧州市池塘养殖凡纳滨对虾为研究对象, 通过持续追踪对虾养殖系统中环境因子、微生物因子和对虾健康指标三要素, 探索池塘养殖对虾AHPND的发生与病原、环境和机体免疫因子三者间的耦合关系, 掌握环境因子及其变化与对虾疾病发生的相互关系, 为池塘养殖对虾疾病临床防控提供依据与技术支撑。
1. 材料与方法
1.1 养殖背景与样品采集
本实验在河北省沧州市对虾池塘养殖基地进行, 养殖用水为渤海水源, 养殖区共有水面133 km2, 全部为池塘养殖模式, 单个池塘面积为0.02—0.04 km2不等, 平均有效水深1.4 m, 每年4月中旬至5月下旬集中投苗, 投苗密度约为25—35尾/m2。本试验共选择核心养殖区内4个无串联的池塘作为试验点(分别标记为Pond 1、Pond 2、Pond 3和Pond 4), 养殖池塘基本信息见表 1, 试验池塘所处位置具有独立的进排水系统, 在保证生产管理方式统一的情况下, 试验池塘间无水质交换和生物污染。全程投喂商业化配合饵料, 投喂频率为2次/日, 投饵量为对虾体重的3%。并利用市售商业化AHPND检测试剂盒(DHelix, 031092M Ⅱ)定期监测池塘发病风险和确定疾病类型。
表 1 试验池塘基本信息Table 1. Basic information of the four test ponds编号
No.面积
Aera (hm2)水深
Depth (m)苗种投放密度
Density of initial
shrimp postlarva
(ind./m2)养殖时间
Breeding time (d)投喂时间
Feeding timePond 1 2.40 1.50 22.5 68 5:00; 16:30 Pond 2 3.33 1.30 35.0 61 6:30; 16:30 Pond 3 3.33 1.30 26.0 70 5:00; 16:30 Pond 4 3.20 1.30 21.5 60 5:30; 16:40 本实验自2020年7月5日开始, 8月8日结束, 共35d, 每天早上7点开始进行样品采集工作: (1)观察记录各试验点对虾的体色、活力、摄食强度、健康程度和生产管理情况, 并记录气候、气温、风力、降雨量等气候因子; (2)通过定点撒网随机捕捞10尾对虾装入含原池水的采样箱中, 打包充气低温带回基地实验室进行可培养微生物和免疫酶活性分析; (3)利用采水器采集各池塘离岸2 m、水下0.5 m处的水样1.5 L, 低温带回基地实验室进行可培养微生物与氨氮(NH4-N)/亚硝态氮(NO2-N)检测。
1.2 水质理化因子测定
每个实验池塘设置3个不同采样点, 采用YSI便携式水质测定仪(proplus, 美国)每日定点测量各实验池塘3个不同位点水体的实时温度、盐度、DO和pH, 并计算各池塘内3个试验点均值。取100 mL水样利用0.45 μm滤膜进行抽滤处理, 取过滤后的海水测定水体氨氮和亚硝态氮含量, 测量方式采用WTW 6600分光光度仪结合氨氮、亚硝态氮检测试剂盒(Supelco, ISO7150和ISO6777), 每组样品平行测定3次并计算均值。
1.3 虾体和水体中可培养细菌和弧菌数量分析
每个试验池塘随机剪取5尾对虾肝胰腺组织, 每尾对虾取约0.2 g, 混合后称重并加入0.5 mL无菌PBS溶液进行匀浆, 用无菌1.5% NaCl溶液对匀浆液进行10倍梯度稀释, 取稀释后的溶液100 μL涂布于TSB(胰蛋白胨大豆肉汤)和TCBS(硫代硫酸盐柠檬酸盐胆盐蔗糖)琼脂培养基上, 28℃培养24h后进行菌落计数。根据特定稀释梯度下TSB和TCBS平板上所生长的菌落数量推算单位虾体肝胰腺内可培养细菌与弧菌总数。同步对池塘水样进行梯度稀释和可培养细菌与弧菌数量统计。
1.4 可培养优势细菌的鉴定
根据TSB和TCBS琼脂培养基上的细菌形态、大小及颜色区分优势细菌种类和数量, 随后, 挑取优势单菌落接种于新的TSB琼脂培养基上进行优势菌株的分离纯化, 在28℃培养24h后用灭菌枪头刮取纯化后的优势菌保存于含20%甘油的保种液中, –80℃冰箱冻存。同步挑取优势菌单菌落重悬于100 µL无菌去离子水内并吹吸均匀, 经99℃金属浴15min后, 置于4℃, 12000 r/min的条件下离心2min, 吸取上清液作为PCR扩增的DNA模板, 采用分光光度法测定DNA浓度并用无菌超纯水将其调整为约100 ng/μL, 用作PCR模板DNA, 并进行16S rDNA基因序列扩增。上下游引物分别为5′-GAGTTTGATCCTGGCTCAG-3′ (27F)和5′-GGTTACCTTGTTACGACTT-3′ (1492R)[16]。PCR反应体系(50 µL)为: 2×Master Mix 25.0 µL, 上下游引物各1.0 µL, 模板DNA 1.0 µL, dH2O 22.0 µL。扩增条件为: 94℃预变性5min; 94℃变性30s、52.5℃退火30s、72℃延伸1.5min, 循环30次; 72℃温育10min, 4℃保存。PCR产物通过1%琼脂糖凝胶电泳检测合格后委托生工生物工程(上海)股份有限公司进行测序, 所得序列在BLAST数据库和EZBioCloud标准菌株数据库进行同源比对, 对鉴定出的优势菌株种类和数量进行统计分析。
1.5 对虾肌肉免疫酶活性测定
用75%酒精棉擦拭对虾体表, 待酒精挥发后剪取尾部肌肉, 每个池塘5尾虾混为一个样品并称重约1.0 g, 等比加入0.9%无菌NaCl溶液进行混合, 并置于冰水浴条件下进行匀浆, 匀浆后在4℃、4000 r/min条件下离心10min后取上清液[17]。使用南京建成生物技术有限公司的免疫酶活试剂盒测定肌肉内的免疫酶活性, 检测方法按照试剂盒说明书进行。检测指标主要包括碱性磷酸酶(AKP)、酸性磷酸酶(ACP)、溶菌酶(LZM)、超氧化物歧化酶(SOD)和酚氧化酶(PO)等5种对虾非特异性免疫酶活性。
2. 结果
2.1 池塘养殖凡纳滨对虾健康监测及管理信息
实验期间试验区多为晴朗天气, 其中8场降雨分别出现在7月6日(5.33 mm)、7月11日(19.6 mm)、7月12日(1.57 mm)、7月13日(47 mm)、7月17日(40 mm)、7月26日(47 mm)、7月31日(110 mm)和8月1日(47 mm) (降雨量数据来源: 中国气象网)。试验期内各池塘发病情况不一, 除Pond 1以外的3个试验点在观察期不同时间均出现一定的肝胰腺坏死症状, 各池塘采样对虾出现典型AHPND发病症状且伴有对虾死亡的时间分布见表 2。Pond 2和Pond 4试验点发病较严重, 死亡率较高, 期间采取泼洒复方中草药制剂和微生态制剂进行治疗与疾病防控, Pond 3试验点对虾两次发病病程较短, 采用大量换水处理后症状消失。
表 2 各实验点对虾出现典型AHPND发病和死亡时的信息统计Table 2. AHPND occurrence and distribution information in each test sites实验点
Groups对虾AHPND检测阳性且具有典型肝胰腺坏死症状
发生时间Time points of AHPND positive with typical hepatopancreas necrosis symptom7.7 7.10 7.12 7.15 7.16 7.26 Pond 1 Pond 2 + + + Pond 3 + + Pond 4 + + + 注: “+”代表发病, 空白代表未发病Note: The “+” means outbeak of AHPND and the blank means absence points 2.2 环境因子监测及其与对虾健康状况的相关性分析
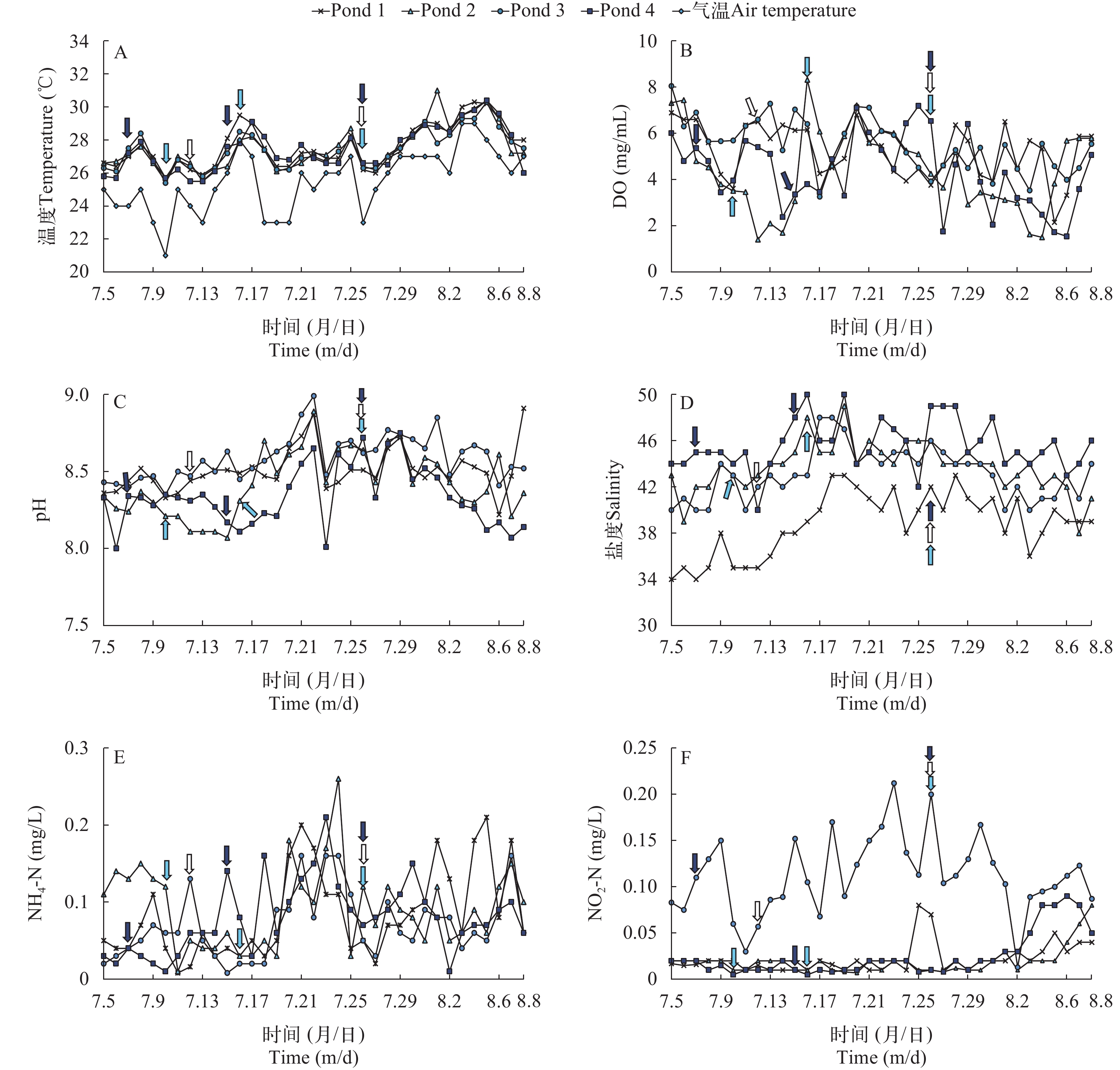
温度 统计分析表明, 在试验周期内, 对虾养殖池塘范围内, 离岸气温整体呈先升高后下降趋势, 清晨气温最高29.0℃, 最低21.0℃, 平均气温24.6℃。实验池塘水温变化与离岸气温呈相似的波动趋势, 波动范围为24.8—31.0℃, 随着时间的增长呈波动上升, 且水温的变化与气温同步但相对缓和(图 1A)。
![]() 图 1 实验周期内池塘水质因子变化趋势A. 温度; B. DO; C. pH; D. 盐度; E. 氨氮; F. 亚硝态氮;
图 1 实验周期内池塘水质因子变化趋势A. 温度; B. DO; C. pH; D. 盐度; E. 氨氮; F. 亚硝态氮;. pond 2 发病节点; . pond 3 发病节点; . pond 4 发病节点 Figure 1. Trends of environmental factors in the ponds water during the experiment periodA. water and air temperature; B. DO; C. pH; D. salinity; E. NH4-N; F. NO2-N;. Time of AHPND occurrence in pond 2; . Time of AHPND occurrence in pond 3; . Time of AHPND occurrence in pond 4 DO 孔巧香等[18]认为南美白对虾养殖过程中良好DO水平在4.00 mg/L以上。试验周期内4个试验点DO变化明显, 波动范围为1.4—8.32 mg/L, Pond 1和Pond 3试验点基本保持在4.00 mg/L以上, Pond 2和Pond 4试验点有超过40%的天数处于4.00 mg/L以下(图 1B)。
pH 试验期内pH整体变化幅度较小, 波动范围为8.0—8.91, Pond 1和Pond 3试验点呈现先增高后降低的变化趋势, Pond 2和Pond 4试验点呈先降低后增高再降低趋势。140次pH检测数据中有5次超过8.8, 12次低于8.2, 大部分处于8.2—8.8, 为对虾健康养殖水质pH的正常波动范围(图 1C)。
盐度 试验周期内养殖水体盐度变化范围较小, 整体处于34—50, 平均盐度为42.82, 盐度范围较常规海水盐度偏高, 通过地理区域分析表明, 盐度变化受地理区系、海区海水盐度、换水量、降雨和水分蒸发等多种因素共同影响(图 1D)。
氨氮和亚硝态氮 试验周期内各养殖池水体中氨氮和亚硝态氮浓度均较低。氨氮整体变化幅度在0.01—0.26 mg/L, 且随着监测时间的推移整体略有增高趋势; 亚硝态氮监测初期含量较低, 整个周期变化相对平稳, 在监测后期略有增加的趋势, 变化幅度为0.01—0.21 mg/L(图 1E, F)。
结合试验周期内池塘对虾健康状况分析表明, 患病池塘出现典型AHPND发病症状和死亡前1—2d, 水体温度先上升达到峰值后再下降; DO随着水温的增高降幅滞后明显, 且与天气阴晴密切相关; pH随着温度的增高小幅度增高, 降雨后小幅度下降; 4个试验点, 氨氮和亚硝态氮随着水温升高也呈一定升高趋势, 但均处于较低水平, 与疾病发生关联性较低。
2.3 对虾养殖池塘水体和虾体内可培养微生物变化规律及其与对虾健康水平的相关性分析
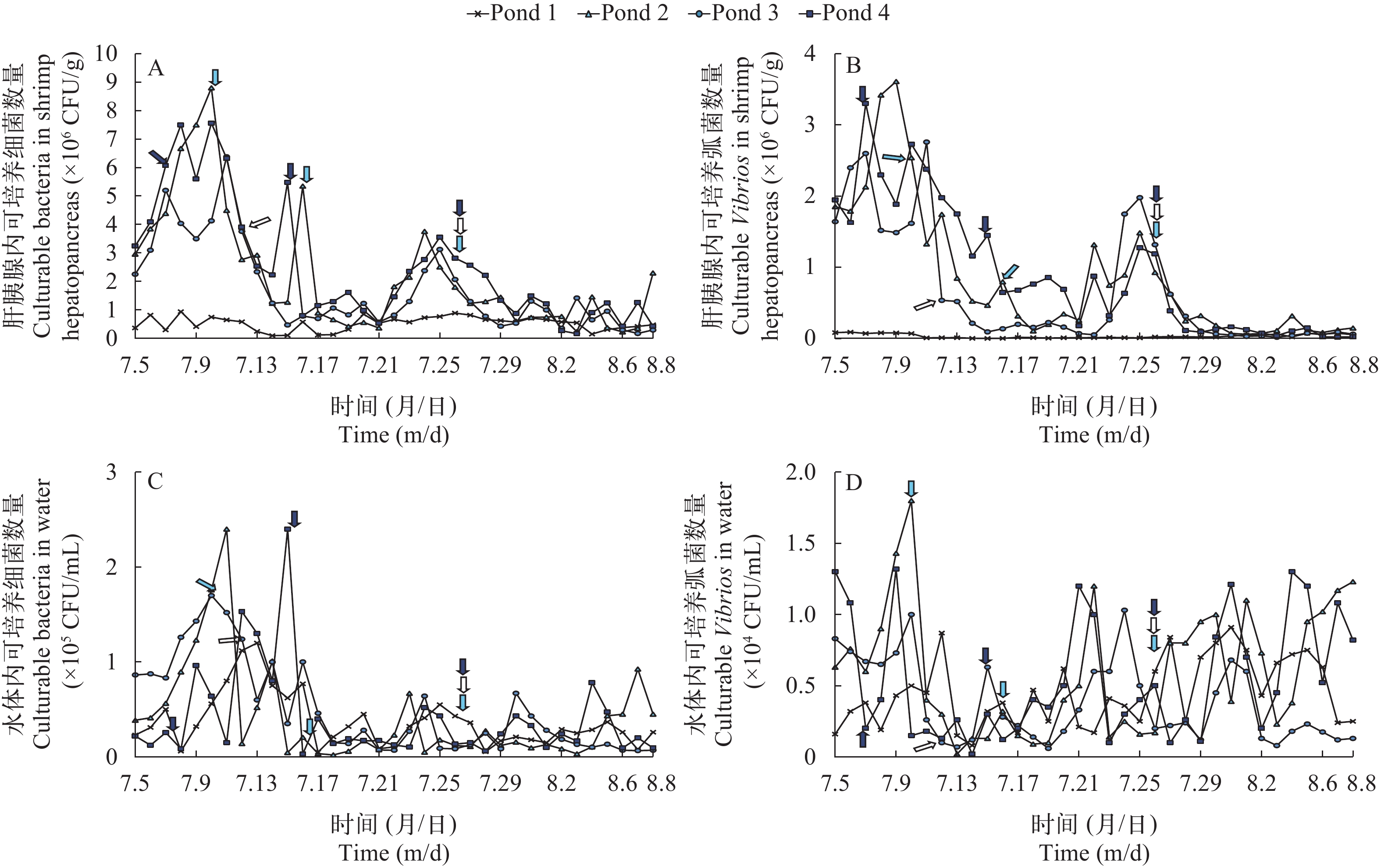
通过对试验周期内各试验点虾体和水体中可培养微生物含量进行统计分析表明, Pond 2、Pond 3和Pond 4试验点虾体肝胰腺内可培养微生物数量大多处于106 CFU/g, 水体中细菌和弧菌数量分别处于104—105和103—104 CFU/mL数量级范围内。Pond 1试验点虾体细菌和弧菌数量稳定维持在105和104 CFU/g以下, 水体内细菌和弧菌数量稳定维持在104和103 CFU/mL以下。Pond 1试验点内虾体和水体可培养微生物数量较其他3个池塘低至少1个数量级(图 2A—D)。
![]() 图 2 实验周期内池塘水体和对虾肝胰腺内细菌和弧菌数量变化趋势A. 虾体内细菌数量; B. 虾体内弧菌数量; C. 水体细菌数量; D. 水体弧菌数量;
图 2 实验周期内池塘水体和对虾肝胰腺内细菌和弧菌数量变化趋势A. 虾体内细菌数量; B. 虾体内弧菌数量; C. 水体细菌数量; D. 水体弧菌数量;. pond 2 发病节点; . pond 3 发病节点; . pond 4 发病节点 Figure 2. Trend of the number of culturable bacteria in water and shrimp hepatopancreas during the experiment periodA. the number of culturable bacteria in shrimp hepatopancreas; B. the number of culturable Vibrios in shrimp hepatopancreas; C. the number of culturable bacteria in water; D. the number of culturable Vibrios in water;. time of AHPND occurrence in pond 2; . time of AHPND occurrence in pond 3; . time of AHPND occurrence in pond 4 结合对虾健康状况分析表明, 对虾出现典型AHPND临床症状期间, 虾体细菌和弧菌数量处于106和105 CFU/g水平, 在出现症状前1—2d达到峰值, 且多伴随水温升高和DO的降低; 水体中可培养细菌和弧菌数量变化较大, 且虾体细菌和弧菌数量与水体相比存在明显的滞后效应。Pond 1试验点全程未发病, 虽然细菌数在105 CFU/g水平波动, 但弧菌量整体较低。比较分析表明, 对虾患病越严重的池塘内弧菌数量越高。
2.4 池塘养殖凡纳滨对虾肝胰腺中可培养优势菌鉴定
检测期间共鉴定优势菌株219株, 分别包含弧菌属(Vibrio)、发光杆菌属(Photobacterium)、希瓦氏菌属(Shewanella)、芽孢杆菌属(Bacillus)、假交替单胞菌属(Pseudoalteromonas)、假单胞菌属(Pseudomonas)、不动杆菌属(Acinetobacter)、气单胞菌属(Aeromonas)和微小杆菌属(Exiguobacterium)等9个属, 共41个种(表 3)。弧菌属菌株数量共135株, 19种, 占全部鉴定菌株的61.6%, 其中包含欧文氏弧菌(V. owensii)、坎贝氏弧菌(V. campbellii)、副溶血弧菌(V. parahaemolyticus)、溶藻弧菌(V. alginolyticus)和哈维氏弧菌(V. harveyi)等与AHPND发生相关的菌株99株, 占总弧菌数的73.3%。此外, 芽孢杆菌属菌株数量达到38株, 占全部鉴定菌株的17.4%, 结合养殖管理分析, 芽孢杆菌作为优势菌检出的日期主要集中在使用微生态制剂后的1—2d。
表 3 基于16S rDNA的对虾肝胰腺可培养优势菌株鉴定信息Table 3. The composition of dominant bacteria in the L. vannamei pond culture system菌株
Strain数量
Number菌株
Strain数量
NumberVobrio弧菌属 135 Bacillus芽孢杆菌属 38 欧文氏弧菌V. owensii 41 枯草芽孢杆菌B. subtilis 17 坎贝氏弧菌V. campbellii 23 暹罗芽孢杆菌B. siamensis 10 溶藻弧菌V. alginolyticus 21 特基拉芽孢杆菌B. tequilensis 4 副溶血弧菌V. parahaemolyticus 10 弯曲芽孢杆菌B. flexus 2 肝弧菌V. hepatarius 8 短小芽孢杆菌B. pumilus 2 需钠弧菌V. natriegens 6 黄海芽胞杆菌B.marisflavi 2 V. hyugaensis 5 B. cabrialesii 1 哈维氏弧菌V. harveyi 4 Photobacterium发光杆菌属 26 轮虫弧菌V. rotiferianus 3 美人鱼发光杆菌P. damselae 25 蓝弧菌V. azureus 2 海洋发光杆菌P. marinum 1 日本弧菌V. japonicus 2 Exiguobacterium微小杆菌属 6 沙蚕弧菌V. nereis 2 墨西哥微小杆菌E. mexicanum 3 变异弧菌V. variabilis 2 印度微小杆菌E. indicum 2 锡那罗州弧菌V. sinaloensis 1 深海微小杆菌E. profundum 1 V. ruber 1 Acinetobacter不动杆菌属 5 萨迦米弧菌V. sagamiensis 1 约氏不动杆菌A. johnsonii 2 拟态弧菌V. mimicus 1 琼氏不动杆菌A. junii 2 非典型弧菌V. atypicus 1 皮特不动杆菌A. pittii 1 巴西弧菌V. brasiliensis 1 Pseudomonas假单胞菌属 2 Shewanella希瓦氏菌属 3 P. oryzihabitans 1 海滨希瓦氏菌S. litorisediminis 2 恶臭假单胞菌P. alloputida 1 瓦氏希瓦氏菌S. waksmanii 1 维氏气单胞菌Aeromonas veronii 2 假交替单胞菌 Pseudoalteromonas shioyasakiensis 2 2.5 对虾机体免疫酶活及其与对虾健康状况相关性分析
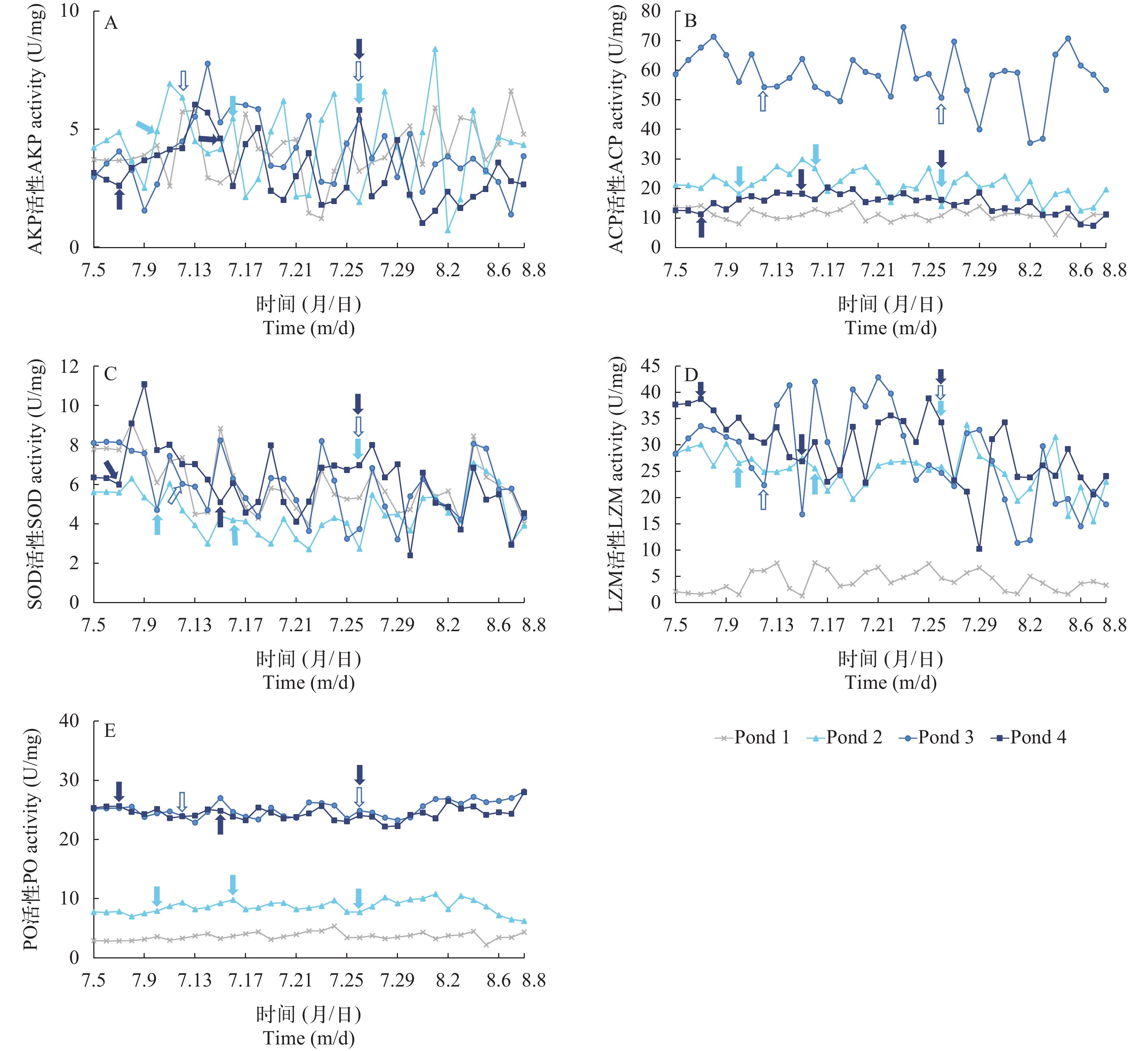
通过对试验周期内各试验点对虾体内免疫酶活性进行统计分析表明, ACP、LZM、AKP和SOD整体波动明显, PO变化幅度较小(图 3)。AKP和SOD的活性在各组内均波动明显, 活性变化范围为1.0—8.5和2.4—11.07 U/mg, 且无显著的组间差异和疾病发生相关性。Pond 3试验点ACP活性最高, 波动范围在35.4—74.5 U/mg, 其后依次为Pond 2和Pond 4试验点, ACP活性变化范围为12.6—29.9和7.4—20.3 U/mg, Pond 1试验点活性总体保持在最低, 波动范围为4.4—15.2 U/mg; Pond 1试验点LZM活性在1.3—7.6 U/mg, 其他3个试验点在10.2—42.8 U/mg, 且出现典型肝胰腺坏死症状时LZM活性均高于25.6 U/mg; Pond 3和Pond 4试验点PO活性最高, 波动范围在22.2—28.1 U/mg, Pond 2试验点PO活性变化范围为6.2—10.8 U/mg, Pond 1试验点活性最低, PO活性变化范围为2.2—5.4 U/mg。
![]() 图 3 四个实验点监测周期内对虾机体门免疫酶活性的变化趋势A. AKP; B. ACP; C. SOD; D. LZM; E. PO;
图 3 四个实验点监测周期内对虾机体门免疫酶活性的变化趋势A. AKP; B. ACP; C. SOD; D. LZM; E. PO;. pond 2 发病节点; . pond 3 发病节点; . pond 4 发病节点 Figure 3. Four experimental points monitor the trend of changes in shrimp phyla immune enzyme activity during the periodA. AKP; B. ACP; C. SOD; D. LZM; E. PO;. Time of AHPND occurrence in pond 2; . Time of AHPND occurrence in pond 3; . Time of AHPND occurrence in pond 4 结合对虾健康状况分析表明, 发病池塘与未发病池塘内对虾肌肉中AKP和SOD活性波动明显但无明显组间差异性; 检测期间Pond 1试验点全程处于健康状态, 且该实验组对虾肌肉内ACP、LZM、PO活性水平全程较低, Pond 3试验点的对虾疾病暴发迅猛, 以上3种免疫酶活性水平整体升高, 且发病池塘Pond 2和Pond 4对虾肌肉中的LZM和PO活性全程高于Pond 1试验点, 综合分析表明, 低水平的LZM和PO可作为池塘养殖系统内对虾健康状况的潜在指示因子。
2.6 监测因子权重分析
在极差、均值、标准差和方差等统计量中, 标准差是最常用、最重要的离散趋势统计量(表 4), 标准差越大, 表示变量值之间的差异越大, 各数据距离均值越远, 则平均数的代表性就越低。反之, 标准差越小, 表示变量值之间的差异越小, 各数据距离均值较近, 则平均数的代表性就越高。比较表 2中各水质理化因子的描述统计量, 按标准差从大到小依次排列为盐度>DO>温度>pH>亚硝态氮>氨氮, 由此可知在本次监测过程中, 盐度、DO和温度变化幅度比较大, pH、亚硝态氮和氨氮变化幅度较小即在池塘养殖模式下最易控制; 比较表 2中各虾体免疫因子的描述统计量, 按标准差从大到小依次排列为ACP>LZM>SOD>AKP>PO, 可知在养殖过程中, ACP、LZM变化幅度比较大, SOD、AKP和PO变化幅度比较小; 可培养细菌标准差由大到小依次为虾体细菌>虾体弧菌>水体细菌>水体弧菌, 虾体可培养细菌和胡菌含量变化幅度较大。
表 4 各理化因子的描述统计量Table 4. Descriptive statistics of each physical and chemical factor统计量
Statistic全距
Full range极大值
Maximum极小值
Minimum均值
Average value标准差
Standard deviation方差
Variance水温Water temperature (℃) 35 4.9 30.3 25.4 27.4 1.132 1.28 DO (mg/L) 35 4.79 5.93 4.46 5.5 1.15 1.32 盐度Salinity (‰) 35 8 48 40 43 2.21 4.9 pH 35 0.64 8.99 8.35 8.59 0.14 0.02 氨氮NH4-N (mg/L) 35 0.15 0.16 0.01 0.07 0.042 0.0018 亚硝态氮NO2-N (mg/L) 35 0.199 0.212 0.013 0.12 0.044 0.002 SOD 35 5 8.23 3.22 5.94 1.53 2.35 PO 35 5.27 28.12 22.85 25.14 1.33 1.76 AKP 35 10.35 12.53 2.15 4.23 1.89 1.83 ACP 35 41.3 74.55 33.32 5.81 8.86 76 LZM 35 37.25 42.84 5.59 27 9.69 72 水体细菌
Culturable bacteria in water35 1.79×105 1.86×105 7000 6.02×107 4.72×104 2.23×109 水体弧菌
Culturable Vibrios in water35 1.37×105 1.38×105 800 3.08×107 3.38×104 1.14×109 虾体细菌
Culturable bacteria in shrimp hepatopancreas35 7.88×107 7.90×107 1.76×105 4.54×1012 1.30×107 1.68×1014 虾体弧菌
Culturable Vibrios in shrimp hepatopancreas35 4.47×106 4.52×106 5.50×104 2.88×1010 1.03×106 1.07×1012 3. 讨论
系统解析病原、宿主和环境间的耦合关系, 是建立和发展水生动物疫病防控策略的核心[19]。本文通过对池塘养殖模式下凡纳滨对虾AHPND发生及其环境、病原、虾体免疫因子进行持续性跟踪监测, 系统解析了AHPND发生与环境、病原和虾体免疫间的相互关系。研究表明, 环境因子、微生物因子和机体免疫因子之间存在相互作用, 且对于池塘养殖对虾AHPND发生都有一定的指示作用, 其中温度、DO、可培养细菌和弧菌数量、LZM和PO等指示因子与对虾健康具有直接相关性。
环境因子作为养殖生态系统的重要组成部分, 对养殖生物的存活、生长及生理代谢均有很明显的影响[20]。研究表明, 降雨会改变水体的盐度、水温、pH和DO等, 且一般雨后的1—2d内容易暴发病害[21]。这与本文监测结果较为一致。在本试验周期内, 盐度明显受降雨、换水和蒸发的影响, 与AHPND的发病无直接相关性。温度变化会导致水质变化加快, 水质的不稳定容易引起疾病暴发[22]。本文监测结果发现水温受气温和降雨的双重影响, 其改变会引起水体中DO、pH、氨氮和亚硝态氮的联动变化。研究表明凡纳滨对虾养殖池DO含量应不低于3 mg/L, 低溶解氧水平会造成对虾缺氧, 进而增加疾病暴发风险[23]。监测期间对虾发病前后DO可降到1.4 mg/L, DO过低可能是导致对虾发病的重要因素。此外, 有研究认为氨氮和亚硝酸盐的安全浓度为0.5 mg/L以下[24]。但较低浓度的氨氮和亚硝酸盐胁迫也能降低凡纳滨对虾的生理功能和抵抗力, 增加对副溶血弧菌的易感性[25]。本实验所测氨氮浓度最高为0.26 mg/L, 亚硝酸盐浓度最高为0.21 mg/L, 均处在安全浓度范围内, 说明在大水面池塘养殖模式下氨氮和亚硝酸盐对对虾健康的影响较小, 但监测其浓度的变化能够反应对虾的健康状况。
弧菌是对虾养殖中最常见的病原菌, 对虾体内和养殖水体的细菌在对虾病害暴发期间主要以弧菌丰度的增加为主[26, 27]。已有报道发现弧菌是多种对虾疾病的主要病原菌, 弧菌类疾病已经成为虾类养殖的重要病害之一[28, 29]。AHPND作为近年来最主要的弧菌类疾病, 给对虾养殖带来的危害也越来越严重[30]。据报道,该疾病是通过AHPND发生池塘的共生或水介导而引起的[31]。AHPND病原复杂且多元化, 本实验监测到发病前虾体和水体弧菌数量分别达到105 CFU/g和104 CFU/mL以上, 经细菌鉴定主要是以欧文氏弧菌(V. owensii)、坎贝氏弧菌(V. campbellii)、副溶血弧菌(V. parahaemolyticus)、哈维氏弧菌(V. harveyi)和溶藻弧菌(V. alginolyticus)等为主导, 说明监测的对虾肝胰腺坏死症出现是由多种致病弧菌的大量扩增共同引起的。通过本文研究初步表明, 虾体弧菌数在104 CFU/g, 水体弧菌数在103 CFU/mL以下可作为池塘养殖对虾可培养细菌数量的安全监测范围。
由于对虾缺乏抗体介导的特异性免疫反应, AKP、LZM、ACP、SOD和PO等非特异性免疫酶活性常被用作衡量凡纳滨对虾免疫能力的重要指标[32-34]。研究表明, PO、LZM、SOD和ACP等免疫酶活性会在氨态氮、亚硝态氮、温度、盐度和pH等环境因子的调控下下降[35-37], LZM对水温尤其敏感, 温度的骤升骤降往往会影响酶的活性[38, 39]。病原菌的侵入会引起ACP、LZM活性升高, 发病结束后活性下降。本研究表明, 各试验点对虾肌肉内PO活性稳定维持在一个相对高的水平, 不随外界干扰变化。Pond 1试验点未发病且ACP、LZM和PO活性最低, Pond 3试验点发病病程短、结束快, 3种免疫酶活性水平整体最高, 表明外界刺激会引起免疫酶活性的升高, 自身免疫应激水平越高抵抗疾病的能力越强。
综上所述, 本文系统性完成了凡纳滨对虾池塘养殖系统内的环境因子、微生物因子和健康指示因子与对虾AHPND发生的关联分析, 初步掌握AHPND发生与各因子的相互关系。并选定温度、DO、可培养细菌和弧菌数量、LZM、PO等指示因子作为从“环境-病原-宿主”三方面预警池塘养殖凡纳滨对虾AHPND发生的候选警示因子。相关研究结果为进一步构建对虾疾病发生预警模型及池塘养殖凡纳滨对虾疾病临床防控提供数据支撑。
-
![]()
图 1 实验周期内池塘水质因子变化趋势
A. 温度; B. DO; C. pH; D. 盐度; E. 氨氮; F. 亚硝态氮;
. pond 2 发病节点; . pond 3 发病节点; . pond 4 发病节点 Figure 1. Trends of environmental factors in the ponds water during the experiment period
A. water and air temperature; B. DO; C. pH; D. salinity; E. NH4-N; F. NO2-N;
. Time of AHPND occurrence in pond 2; . Time of AHPND occurrence in pond 3; . Time of AHPND occurrence in pond 4 ![]()
图 2 实验周期内池塘水体和对虾肝胰腺内细菌和弧菌数量变化趋势
A. 虾体内细菌数量; B. 虾体内弧菌数量; C. 水体细菌数量; D. 水体弧菌数量;
. pond 2 发病节点; . pond 3 发病节点; . pond 4 发病节点 Figure 2. Trend of the number of culturable bacteria in water and shrimp hepatopancreas during the experiment period
A. the number of culturable bacteria in shrimp hepatopancreas; B. the number of culturable Vibrios in shrimp hepatopancreas; C. the number of culturable bacteria in water; D. the number of culturable Vibrios in water;
. time of AHPND occurrence in pond 2; . time of AHPND occurrence in pond 3; . time of AHPND occurrence in pond 4 ![]()
图 3 四个实验点监测周期内对虾机体门免疫酶活性的变化趋势
A. AKP; B. ACP; C. SOD; D. LZM; E. PO;
. pond 2 发病节点; . pond 3 发病节点; . pond 4 发病节点 Figure 3. Four experimental points monitor the trend of changes in shrimp phyla immune enzyme activity during the period
A. AKP; B. ACP; C. SOD; D. LZM; E. PO;
. Time of AHPND occurrence in pond 2; . Time of AHPND occurrence in pond 3; . Time of AHPND occurrence in pond 4 表 1 试验池塘基本信息
Table 1 Basic information of the four test ponds
编号
No.面积
Aera (hm2)水深
Depth (m)苗种投放密度
Density of initial
shrimp postlarva
(ind./m2)养殖时间
Breeding time (d)投喂时间
Feeding timePond 1 2.40 1.50 22.5 68 5:00; 16:30 Pond 2 3.33 1.30 35.0 61 6:30; 16:30 Pond 3 3.33 1.30 26.0 70 5:00; 16:30 Pond 4 3.20 1.30 21.5 60 5:30; 16:40  下载: 导出CSV
下载: 导出CSV
表 2 各实验点对虾出现典型AHPND发病和死亡时的信息统计
Table 2 AHPND occurrence and distribution information in each test sites
实验点
Groups对虾AHPND检测阳性且具有典型肝胰腺坏死症状
发生时间Time points of AHPND positive with typical hepatopancreas necrosis symptom7.7 7.10 7.12 7.15 7.16 7.26 Pond 1 Pond 2 + + + Pond 3 + + Pond 4 + + + 注: “+”代表发病, 空白代表未发病Note: The “+” means outbeak of AHPND and the blank means absence points
下载: 导出CSV
表 3 基于16S rDNA的对虾肝胰腺可培养优势菌株鉴定信息
Table 3 The composition of dominant bacteria in the L. vannamei pond culture system
菌株
Strain数量
Number菌株
Strain数量
NumberVobrio弧菌属 135 Bacillus芽孢杆菌属 38 欧文氏弧菌V. owensii 41 枯草芽孢杆菌B. subtilis 17 坎贝氏弧菌V. campbellii 23 暹罗芽孢杆菌B. siamensis 10 溶藻弧菌V. alginolyticus 21 特基拉芽孢杆菌B. tequilensis 4 副溶血弧菌V. parahaemolyticus 10 弯曲芽孢杆菌B. flexus 2 肝弧菌V. hepatarius 8 短小芽孢杆菌B. pumilus 2 需钠弧菌V. natriegens 6 黄海芽胞杆菌B.marisflavi 2 V. hyugaensis 5 B. cabrialesii 1 哈维氏弧菌V. harveyi 4 Photobacterium发光杆菌属 26 轮虫弧菌V. rotiferianus 3 美人鱼发光杆菌P. damselae 25 蓝弧菌V. azureus 2 海洋发光杆菌P. marinum 1 日本弧菌V. japonicus 2 Exiguobacterium微小杆菌属 6 沙蚕弧菌V. nereis 2 墨西哥微小杆菌E. mexicanum 3 变异弧菌V. variabilis 2 印度微小杆菌E. indicum 2 锡那罗州弧菌V. sinaloensis 1 深海微小杆菌E. profundum 1 V. ruber 1 Acinetobacter不动杆菌属 5 萨迦米弧菌V. sagamiensis 1 约氏不动杆菌A. johnsonii 2 拟态弧菌V. mimicus 1 琼氏不动杆菌A. junii 2 非典型弧菌V. atypicus 1 皮特不动杆菌A. pittii 1 巴西弧菌V. brasiliensis 1 Pseudomonas假单胞菌属 2 Shewanella希瓦氏菌属 3 P. oryzihabitans 1 海滨希瓦氏菌S. litorisediminis 2 恶臭假单胞菌P. alloputida 1 瓦氏希瓦氏菌S. waksmanii 1 维氏气单胞菌Aeromonas veronii 2 假交替单胞菌 Pseudoalteromonas shioyasakiensis 2
下载: 导出CSV
表 4 各理化因子的描述统计量
Table 4 Descriptive statistics of each physical and chemical factor
统计量
Statistic全距
Full range极大值
Maximum极小值
Minimum均值
Average value标准差
Standard deviation方差
Variance水温Water temperature (℃) 35 4.9 30.3 25.4 27.4 1.132 1.28 DO (mg/L) 35 4.79 5.93 4.46 5.5 1.15 1.32 盐度Salinity (‰) 35 8 48 40 43 2.21 4.9 pH 35 0.64 8.99 8.35 8.59 0.14 0.02 氨氮NH4-N (mg/L) 35 0.15 0.16 0.01 0.07 0.042 0.0018 亚硝态氮NO2-N (mg/L) 35 0.199 0.212 0.013 0.12 0.044 0.002 SOD 35 5 8.23 3.22 5.94 1.53 2.35 PO 35 5.27 28.12 22.85 25.14 1.33 1.76 AKP 35 10.35 12.53 2.15 4.23 1.89 1.83 ACP 35 41.3 74.55 33.32 5.81 8.86 76 LZM 35 37.25 42.84 5.59 27 9.69 72 水体细菌
Culturable bacteria in water35 1.79×105 1.86×105 7000 6.02×107 4.72×104 2.23×109 水体弧菌
Culturable Vibrios in water35 1.37×105 1.38×105 800 3.08×107 3.38×104 1.14×109 虾体细菌
Culturable bacteria in shrimp hepatopancreas35 7.88×107 7.90×107 1.76×105 4.54×1012 1.30×107 1.68×1014 虾体弧菌
Culturable Vibrios in shrimp hepatopancreas35 4.47×106 4.52×106 5.50×104 2.88×1010 1.03×106 1.07×1012
下载: 导出CSV
-
[1] Tran L, Nunan L, Redman R M, et al. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp [J]. Diseases of Aquatic Organisms, 2013, 105(1): 45-55. doi: 10.3354/dao02621
[2] Soto-Rodriguez S A, Gomez-Gil B, Lozano-Olvera R, et al. Field and experimental evidence of Vibrio parahaemolyticus as the causative agent of acute hepatopancreatic necrosis disease of cultured shrimp (Litopenaeus vannamei) in Northwestern Mexico [J]. Applied and Environmental Microbiology, 2015, 81(5): 1689-1699. doi: 10.1128/AEM.03610-14
[3] Zhang Q, Liu Q, Liu S, et al. A new nodavirus is associated with covert mortality disease of shrimp [J]. The Journal of General Virology, 2014, 95(12): 2700-2709. doi: 10.1099/vir.0.070078-0
[4] 李吉云, 沈辉, 孟庆国, 等. 对虾急性肝胰腺坏死病(AHPND)流行病学、诊断方法及防控措施的研究进展 [J]. 海洋科学, 2021, 45(3): 163-172. Li J Y, Shen H, Meng Q G, et al. Research progress on the epidemiology, diagnosis, prevention, and control of acute hepatopancreas necrosis in shrimp [J]. Marine Sciences, 2021, 45(3): 163-172.
[5] Lai H C, Ng T H, Ando M, et al. Pathogenesis of acute hepatopancreatic necrosis disease (AHPND) in shrimp [J]. Fish & Shellfish Immunology, 2015, 47(2): 1006-1014.
[6] Kondo H, Van P T, Dang L T, et al. Draft genome sequence of non-Vibrio parahaemolyticus acute hepatopancreatic necrosis disease strain KC13.17. 5, isolated from diseased shrimp in Vietnam [J]. Genome Announcements, 2015, 3(5): e00978-e00915.
[7] Liu P C, Lee K K, Yii K C, et al. Isolation of Vibrio harveyi from diseased kuruma prawns Penaeus japonicus [J]. Current Microbiology, 1996, 33(2): 129-132. doi: 10.1007/s002849900087
[8] Liu L, Xiao, J, Xia X, et al. Draft genome sequence of Vibrio owensii strain SH-14, which causes shrimp acute hepatopancreatic necrosis disease [J]. Genome Announcements, 2015, 3(6): e01395-e01315.
[9] Ahn Y S, Piamsomboon P, Tang K F J, et al. Complete genome sequence of acute hepatopancreatic necrosis disease-causing Vibrio campbellii LA16-V1, isolated from Penaeus vannamei cultured in a Latin American country [J]. Genome Announcements, 2017, 5(37): e01011-e01017.
[10] Esteve M, Herrera F C. Hepatopancreatic alterations in Litopenaeus vannamei (Boone, 1939) (Crustacea: Decapoda: Penaeidae) experimentally infected with a Vibrio alginolyticus strain [J]. Journal of Invertebrate Pathology, 2000, 76(1): 1-5. doi: 10.1006/jipa.2000.4951
[11] Lightner D V, Redman R M, Pantoja C R, et al. Early mortality syndrome affects shrimp in Asia [J]. Global Aquaculture Advocate, 2012, 15(1): 40.
[12] Lee C T, Chen I T, Yang Y T, et al. The opportunistic marine pathogen Vibrio parahaemolyticus becomes virulent by acquiring a plasmid that expresses a deadly toxin [J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(34): 10798-10803. doi: 10.1073/pnas.1503129112
[13] Han J E, Tang K F J, Pantoja C R, et al. qPCR assay for detecting and quantifying a virulence plasmid in acute hepatopancreatic necrosis disease (AHPND) due to pathogenic Vibrio parahaemolyticus [J]. Aquaculture, 2015, 442: 12-15. doi: 10.1016/j.aquaculture.2015.02.024
[14] Thakur D P, Lin C K. Water quality and nutrient budget in closed shrimp (Penaeus monodon) culture systems [J]. Aquacultural Engineering, 2003, 27(3): 159-176. doi: 10.1016/S0144-8609(02)00055-9
[15] Boonyawiwat V, Patanasatienkul T, Kasornchandra J, et al. Impact of farm management on expression of early mortality syndrome/acute hepatopancreatic necrosis disease (EMS/AHPND) on penaeid shrimp farms in Thailand [J]. Journal of Fish Diseases, 2017, 40(5): 649-659. doi: 10.1111/jfd.12545
[16] 王凯, 王印庚, 姜勇, 等. 一株感染深水网箱养殖许氏平鲉的病原菌分离与鉴定 [J]. 渔业科学进展, 2019, 40(1): 119-126. Wang K, Wang Y G, Jiang Y, et al. Isolation, identification, and biological characteristics of a pathogenic bacterial strain from cage-cultured black rockfish (Sebastes schlegelii) [J]. Progress in Fishery Sciences, 2019, 40(1): 119-126.
[17] 张喆, 李健, 冯伟, 等. 不同浓度诺氟沙星对中国对虾非特异性免疫酶活的影响 [J]. 渔业科学进展, 2011, 32(2): 53-59. doi: 10.3969/j.issn.1000-7075.2011.02.009 Zhang Z, Li J, Feng W, et al. Effects of norfloxacin on the non-specific immune response of Fenneropenaeus chinensis [J]. Progress in Fishery Sciences, 2011, 32(2): 53-59. doi: 10.3969/j.issn.1000-7075.2011.02.009
[18] 孔巧香. 南美白对虾主要致病微生物安全预警技术研究 [D]. 舟山: 浙江海洋学院, 2013: 15. Kong Q X. Safety early-warning technology of the main pathogenic microorganisms in Penaeus vanname [D]. Zhoushan: Zhejiang Ocean University, 2013: 15.
[19] 宋林生. 海水养殖贝类病害预警预报技术及其应用 [J]. 大连海洋大学学报, 2020, 35(1): 1-9. Song L S. An early warning system for diseases during mollusc mariculture: exploration and utilization [J]. Journal of Dalian Ocean University, 2020, 35(1): 1-9.
[20] 张健龙. 凡纳滨对虾养殖池水质因子及水生生物研究 [D]. 上海: 上海海洋大学, 2016: 1-7. Zhang J L. Research on water quality factors and aquatic organism in Litopenaeus vannamei [D]. Shanghai: Shanghai Ocean University, 2016: 1-7.
[21] 马真. 凡纳滨对虾集约化养殖水质预警模型的研究 [D]. 青岛: 中国海洋大学, 2010: 45. Ma Z. Study on pre-alarming water quality system for intensively cultured Litopenaeus vannamei [D]. Qingdao: Ocean University of China, 2010: 45.
[22] 贾旭颖, 王芳, 王春生, 等. 温度突变和非离子氨对海水和淡水养殖条件下凡纳滨对虾存活的影响 [J]. 中国海洋大学学报(自然科学版), 2013, 43(10): 33-40. Jia X Y, Wang F, Wang C S, et al. Effect of abrupt change in temperature and non-ionic ammonium on survival of Litopenaeus vannamei in seawater and freshwater [J]. Periodical of Ocean University of China, 2013, 43(10): 33-40.
[23] 马建新, 刘爱英, 宋爱芹. 对虾病毒病与化学需氧量相关关系研究 [J]. 海洋科学, 2002, 26(3): 68-71. doi: 10.3969/j.issn.1000-3096.2002.03.020 Ma J X, Liu A Y, Song A Q. A study on the relationship between cod and shrimp viral disease [J]. Marine Sciences, 2002, 26(3): 68-71. doi: 10.3969/j.issn.1000-3096.2002.03.020
[24] 姜令绪, 潘鲁青, 肖国强. 氨氮对凡纳对虾免疫指标的影响 [J]. 中国水产科学, 2004, 11(6): 537-541. doi: 10.3321/j.issn:1005-8737.2004.06.009 Jiang L X, Pan L Q, Xiao G Q. Effects of ammonia-N on immune parameters of white shrimp Litopenaeus vannamei [J]. Journal of Fishery Sciences of China, 2004, 11(6): 537-541. doi: 10.3321/j.issn:1005-8737.2004.06.009
[25] 黄翔鹄, 李长玲, 郑莲, 等. 亚硝酸盐氮对凡纳滨对虾毒性和抗病相关因子影响 [J]. 水生生物学报, 2006, 30(4): 466-471. doi: 10.3321/j.issn:1000-3207.2006.04.016 Huang X H, Li C L, Zheng L, et al. The toxicity of NO2-N on Litopenaeus vannamei and effects of NO2-N on factors relating to the anti-disease ability [J]. Acta Hydrobiologica Sinica, 2006, 30(4): 466-471. doi: 10.3321/j.issn:1000-3207.2006.04.016
[26] Sullam K E, Essinger S D, Lozupone C A, et al. Environmental and ecological factors that shape the gut bacterial communities of fish: a meta-analysis [J]. Molecular Ecology, 2012, 21(13): 3363-3378. doi: 10.1111/j.1365-294X.2012.05552.x
[27] Xiong J, Zhu J, Wang K, et al. The temporal scaling of bacterioplankton composition: high turnover and predictability during shrimp cultivation [J]. Microbial Ecology, 2014, 67(2): 256-264. doi: 10.1007/s00248-013-0336-7
[28] Jayasree L, Janakiram P, Madhavi R. Characterization of Vibrio spp. associated with diseased shrimp from culture ponds of Andhra pradesh (India) [J]. Journal of the World Aquaculture Society, 2006, 37(4): 523-532. doi: 10.1111/j.1749-7345.2006.00066.x
[29] Jiang X, Zhang X, Ren C, et al. Interleukin-2 enhancer binding factor 2 (ILF2) in Pacific white shrimp (Litopenaeus vannamei): alternatively spliced isoforms with different responses in the immune defenses against Vibrio infection [J]. Developmental & Comparative Immunology, 2021(118): 103975.
[30] Kumar B K, Deekshit V K, Raj J R M, et al. Diversity of Vibrio parahaemolyticus associated with disease outbreak among cultured Litopenaeus vannamei (Pacific white shrimp) in India [J]. Aquaculture, 2014(433): 247-251. doi: 10.1016/j.aquaculture.2014.06.016
[31] Joshi J, Srisala J, Truong V H, et al. Variation in Vibrio parahaemolyticus isolates from a single Thai shrimp farm experiencing an outbreak of acute hepatopancreatic necrosis disease (AHPND) [J]. Aquaculture, 2014, 428/429: 297-302. doi: 10.1016/j.aquaculture.2014.03.030
[32] 陈永康, 陈泽恩, 梁武辉, 等. 酵母水解物对低盐胁迫凡纳滨对虾非特异性免疫及抗氧化能力的影响 [J]. 水产学报, 2021, 45(12): 2061-2071. Chen Y K, Chen Z E, Liang W H, et al. Effect of yeast hydrolysate on non-specific immunity and antioxidant ability of Litopenaeus vannamei under low salinity stress [J]. Journal of Fisheries of China, 2021, 45(12): 2061-2071.
[33] Ngo H V T, Huang H T, Lee P T, et al. Effects of Phyllanthus amarus extract on nonspecific immune responses, growth, and resistance to Vibrio alginolyticus in white shrimp Litopenaeus vannamei [J]. Fish & Shellfish Immunology, 2020(107): 1-8.
[34] 赵红霞, 陈冰, 莫文艳, 等. 饲料添加β-1, 3-葡聚糖对凡纳滨对虾生长性能、血清代谢、免疫相关基因表达和抗亚硝酸氮应激能力的影响 [J]. 水生生物学报, 2021, 45(3): 593-600. doi: 10.7541/2021.2020.047 Zhao H, Chen B, Mo W, et al. Effects of dietary β-1, 3-glucan on growth performance, serum metabolite, immune-related gene expression and resistance to nitrite stress in Litopenaeus vannamei [J]. Acta Hydrobiologica Sinica, 2021, 45(3): 593-600. doi: 10.7541/2021.2020.047
[35] Liu C H, Chen J C. Effect of ammonia on the immune response of white shrimp Litopenaeus vannamei and its susceptibility to Vibrio alginolyticus [J]. Fish & Shellfish Immunology, 2004, 16(3): 321-334.
[36] Zhang K, Pan L, Chen W, et al. Effect of using sodium bicarbonate to adjust the pH to different levels on water quality, the growth and the immune response of shrimp Litopenaeus vannamei reared in zero-water exchange biofloc-based culture tanks [J]. Aquaculture Research, 2017, 48(3): 1194-1208. doi: 10.1111/are.12961
[37] Xian J A, Chen X D, Wang A L, et al. Effects of dietary manganese levels on growth, antioxidant defense and immune responses of the juvenile white shrimp, Litopenaeus vannamei, reared in low salinity water [J]. Fish & Shellfish Immunology, 2013, 34(6): 1747.
[38] Tort L, Rotllant J, Liarte C, et al. Effects of temperature decrease on feeding rates, immune indicators and histopathological changes of gilthead sea bream Sparus aurata fed with an experimental diet [J]. Aquaculture, 2004, 229(1/2/3/4): 55-65.
[39] Langston A L, Hoare R, Stefansson M, et al. The effect of temperature on non-specific defence parameters of three strains of juvenile Atlantic halibut (Hippoglossus hippoglossus L.) [J]. Fish & Shellfish Immunology, 2002, 12(1): 61-76.
-
期刊类型引用(5)
1. 董宣,黄倢. 生物安保保障水产养殖绿色高质量发展:回顾与展望. 渔业科学进展. 2025(01): 161-182 .  百度学术
百度学术
2. 安微,林华,郑晶,李丹丹,张婧,徐国锋. WSSV、IHHNV、EHP和NHPB四重荧光定量PCR检测方法的建立. 食品安全导刊. 2024(12): 99-101+106 . 百度学术
3. 王印庚,于永翔,蔡欣欣,张正,王春元,廖梅杰,朱洪洋,李昊. 基于Deep Forest算法的对虾急性肝胰腺坏死病(AHPND)预警数学模型构建. 渔业科学进展. 2024(03): 171-181 . 百度学术
4. 罗家林,古昌辉,沈佳佳,程蔓,韦云勇,叶香尘,贾庆光,李文红,麻艳群. 广西冬闲稻田养殖三种鲤的生长、肌肉营养品质及理化指标比较. 水产学杂志. 2024(05): 118-126 . 百度学术
5. 潘朝阳,袁海宏,梅帅,黄文,邢东旭,胡俊茹,曹俊明,陈冰. 桑叶粉对凡纳滨对虾生长性能、体成分及肌肉品质的影响. 广东畜牧兽医科技. 2024(05): 41-47 . 百度学术
其他类型引用(1)
计量
- 文章访问数: 1764
- HTML全文浏览量: 575
- PDF下载量: 97
- 被引次数: 6