
MORPHOMETRIC STUDY AND COMPARISON OF SCALLOPS (CHLAMYS FARRERI) CULTURED IN DIFFERENT WATER DEPTH
-
摘要: 为了探究远岸深水海区和近岸浅水海区养殖的栉孔扇贝(Chlamys farreri)形态上的差异, 运用传统形态测量学和几何形态测量学的方法, 对来自近岸浅水海区(15 m水深)和远岸深水海区(30 m水深)底栖养殖的2龄栉孔扇贝进行形态测量学研究和比较分析。结果表明: 深水养殖的栉孔扇贝的壳高(SH)、壳长(SL)和壳宽(SW)都极显著大于浅水养殖的栉孔扇贝(P<0.01); 通过界标点(Landmarks)和半界标点(Semi-landmarks)的方法, 及广义普鲁克分析(GPA)、主成分分析(PCA)和典型变量分析(CVA), 结果表明深水养殖的栉孔扇贝的整体壳形尺寸大于浅水养殖的栉孔扇贝, 除去尺寸大小因素后两者的壳耳和壳体扇形边缘存在显著差异。文章为栉孔扇贝深水养殖的选育和增养殖工作提供基于形态学研究的理论依据, 同时比较分析了传统形态测量学和几何形态测量学的原理和分析方法, 为研究生物体形态差异时选择合适的研究方法提供新参考。Abstract: Shell traits play an important role in the identification and delimitation of mollusk species. Determining the growth stage of Pecten is always a challenge in aquaculture, taxonomy, and evolution. In order to explore the morphological differences of the scallops cultured in the offshore deep-water sea area and the inshore shallow water sea area, traditional morphometry and geometrical morphometry methods were used to obtain their morphological differences. We tested the variation in shape indexes between the two groups of different water depth using the Independent-samples T test. For geometric morphometric analysis, we performed generalized procrustes analysis (GPA) on the collected morphological data coordinates of the two scallops. Then we subjected transformation data to principal component analysis (PCA) and canonical variate analysis (CVA). The results of the traditional multivariate morphometric analysis showed that the shell height (SH), shell length (SL) and shell width (SW) of the scallops in the deep water were significantly larger than the scallops in the shallow water (P<0.01); Through the analysis of landmarks and semi-landmarks variations, the results showed that the overall shell size of the scallops in the deep water was larger than that of the scallops in the shallow water and there is a significant difference in the auricles and disc of the two scallops after removing the size factor. This paper not only provides a theoretical basis for morphological research for the deep-water aquaculture of the scallops, but also providing a reference for selecting appropriate methods in the morphometric research.
-
形态测量学(Morphometric)是用于描述生物体和生物体之间由于生长、环境变化或者进化, 发生的形态变化或差异的分析方法。数理生物学家Thompson最早在1917年提出, 通过测量和坐标变换, 能够从数理角度描述分析外形相似的生物体间的形态差异[1]。目前根据发展时间和方法原理, 可以将形态测量学分为传统形态测量学(Traditional morphometric)和几何形态测量学(Geometric morphometric)。
传统形态测量学, 或称多变量形态测量学, 通过统计学分析, 得到其形态上的变化规律或不同生物体间的形态差异。变量一般为生物体上测量的线性距离或线性距离的比值和乘积, 比如长度、宽度、高度、角度、比率及面积等。王旭等[2]采用8个度量特征对3个地理群体的单齿螺(Monodonta labio)的形态差异进行研究, 发现舟山和厦门群体形态最为接近, 大连群体趋异程度最大, 但差异都未达到亚种水平; 牛泓博等[3]同样利用该方法, 通过对辽宁沿海菲律宾蛤仔的多变量形态度量学分析, 揭示了该种贝类不同群体的形态差异。
几何形态测量学利用轮廓线(Outline)或界标点(Landmarks), 来描述生物体的形态或标记特征部位, 得到其形态上的变化规律或不同生物体间的形态差异[4]。标记的位点构成数据分析中的原始变量, 之后映射到数学坐标中采用多变量数据分析的方法获得其差异性。来益同等[5]使用界标点及半界标点对2种纯合体色品系的大瓶螺的壳口和壳顶进行几何形态测量学分析, 结果表明该贝壳的形态变化主要集中在壳口上半部, 且不同体色和性别的大瓶螺存在着显著的形态差异; 陈楠桦等[6]对4种滨螺的几何形态测量学研究, 揭示了这4种滨螺在壳口和螺体上的主要形态差异和它们之间基于形态的亲缘关系远近。
栉孔扇贝(Chlamys farreri)属软体动物门(Mollusca)、双壳纲(Bivalvia)、海扇蛤目(Pectinida)、海扇蛤科(Pectinidae)、栉孔扇贝属(Chlamys), 是我国北方重要的海水经济养殖贝类品种[7]。目前栉孔扇贝主要的养殖区域集中在近岸的15 m水深的浅水海区, 随着养殖产业的不断发展, 浅水海区的利用日渐饱和, 部分区域资源过度利用, 因养殖密度过大导致了水质恶化、养殖污染加剧、病害频发和种质退化等问题[8, 9]。相较而言, 远岸20—40 m水深的深水海区还有较大的资源开发空间, 因其具有可利用场所广阔、与其他产业在场地利用上矛盾小和水交换能力强等优点, 海水养殖从近岸浅水海区向远岸深水海区拓展, 已成为必然趋势[10-12]。
对于海产双壳类来说, 研究壳的形态有助于揭示其生长规律, 是生物学性状研究的主要内容之一, 也是分类学、进化学和渔业资源管理等多个领域的研究要点[13-20]; 同时有大量研究表明, 贝类壳的形态性状与其活体重、总干重、软体部重等经济性状指标具有相关性, 形态指标易于测量且不损伤贝类的软体部, 在人工选育和提高养殖效益等方面具有重要意义[21-27]。
我国目前对深水养殖的栉孔扇贝的研究不多, 主要集中在存活率、免疫特征、固碳速率等方面[28, 29]。王光花等[30]曾做过桑沟湾远岸水域(30 m水深)养殖栉孔扇贝的形态性状与经济性状之间的相关性和多元回归关系, 但并未将深水养殖的栉孔扇贝和浅水养殖的栉孔扇贝的形态作比较与分析。本文拟通过形态测量学(Morphometric)的方法, 统计和分析深水养殖和浅水养殖的栉孔扇贝的形态学上的差异, 为栉孔扇贝拓展深水养殖的科学选育和增养殖等工作提供理论依据。
1. 材料与方法
1.1 材料
2019年6月, 在大连市黑石礁海域的近岸浅水海区(15 m水深, 38°50′35.37″ N, 121°34′55.36″E)和远岸深水海区(30 m水深, 38°45′55.58″N, E121°35′32.14 E″)分别随机选取底栖养殖的壳形完整的2龄栉孔扇贝30只, 运回农业农村部北方海水增养殖重点实验室(大连)暂养3d后进行测量和解剖。
1.2 形态测量方法
传统形态测量 清除贝壳表面的牡蛎、藤壶和石灰虫等附着物后, 使用电子游标卡尺对贝壳的壳高(SH)、壳长(SL)和壳宽(SW)进行测量, 精确到0.01 mm, 测量部位及指标见图 1。
![]() 图 1 栉孔扇贝传统形态测量学指标1. 深水栉孔扇贝; 2. 浅水栉孔扇贝Figure 1. Traditional morphometrical measurements of C. farreri1. The scallops in the deep water; 2. The scallops in the shallow water
图 1 栉孔扇贝传统形态测量学指标1. 深水栉孔扇贝; 2. 浅水栉孔扇贝Figure 1. Traditional morphometrical measurements of C. farreri1. The scallops in the deep water; 2. The scallops in the shallow water几何形态图像获取 解剖扇贝去除软体部后, 使用数码相机(Canon G12)对每个栉孔扇贝的右壳进行拍摄捕获图像, 以采集二维数据。相机固定在翻拍架上, 使用水平仪调整角度将透镜中心轴与拍摄面垂直, 拍摄距离固定在125 mm, 保证不同样本之间具有相同的比例尺, 减小被拍摄物在投影时由于视差产生形变[31]; 扇贝壳置于水平架, 凹面向上, 以垂直于铰合边缘、从壳顶发出的直线为中轴, 使用水平仪确保壳缘平面与拍摄面平行。为了降低操作的偶然误差, 照片的拍摄和后续数字化工作均由一人完成。
数据的标准化处理 本文采用界标点(Landmarks)+半界标点(Semi-landmarks)的方法[32], 使用ImageJ v1.52对2种不同水深的栉孔扇贝的二维图像进行标点和数据采集工作。界标点的选取要求既能反映研究对象的形态差异, 同时在各样本之间又具有同源性, 半界标点是为了更加精细地确定研究对象的整体轮廓。共选取28个点作为标记点, 其中1—8为界标点, 为扇贝的生物学特征点, 9—28为半界标点, 为沿着贝壳边缘轮廓线的半界标点。具体见图 2和表 1。
表 1 栉孔扇贝界标点与半界标点的位置Table 1. The position of landmarks and semi-landmarks on C. farreri标点Landmark and Semi-landmark 对应部位Position 1 壳顶 2 前耳前端 3 足丝孔 4 贝壳右边缘最宽处 5 壳底 6 贝壳左边缘最宽处 7 后耳拐弯处 8 后耳末端 9—13 沿着后耳轮廓线的半界标点 14—18 沿着前耳轮廓线的半界标点 19—23 沿着贝壳右边缘轮廓线的半界标点 24—28 沿着贝壳左边缘轮廓线的半界标点 ![]() 图 2 栉孔扇贝界标点与半界标点的位置Figure 2. Position of the 8 landmarks and 20 semi-landmarks on C. farreri
图 2 栉孔扇贝界标点与半界标点的位置Figure 2. Position of the 8 landmarks and 20 semi-landmarks on C. farreri1.3 数据分析
传统形态测量 使用Excel 2018计算2种不同水深的栉孔扇贝在传统测量方法下的形态学指标的平均值(Mean)、标准差(SD)和变异系数(CV%)等基本统计量; 使用SPSS v23.0对2种不同水深的栉孔扇贝间的各形态学性状进行独立样本T检验(Independent-samples T test), 获得其差异的显著性。
几何形态测量 对采集到的2种不同水深的栉孔扇贝的形态数据坐标进行广义普鲁克分析(Generalized Procrustes Analysis, GPA), 去除在取界标点时由于位置方向和比例变化产生的非形态差异的影响; 然后将经GPA变换后的数据进行主成分分析(Principal Components Analysis, PCA)和典型变量分析(Canonical Variate Analysis, CVA), 前者是分析每个样本几何形态上的差异, 后者是分析2种不同水深的栉孔扇贝间的形态差异。上述数据分析均使用Past v3.24完成[33]。
2. 结果
2.1 传统形态测量
2种不同水深的栉孔扇贝的传统形态测量结果见表 2。深水养殖的栉孔扇贝的壳高、壳长和壳宽的最大值分别为102.96、97.51和33.71 mm, 最小值分别为69.76、65.81和21.47 mm; 浅水养殖的栉孔扇贝的壳高、壳长和壳宽的最大值分别为74.54、70.13和26.46 mm, 最小值分别为65.86、59.67和19.75 mm。
表 2 传统形态测量学的基本统计量Table 2. Basic statistics of traditional morphometry种类Specimen 统计量
Statistic壳高SH (mm) 壳长SL (mm) 壳宽SW (mm) 深水养殖的栉孔扇贝
C. farreri from the deep water最大值 102.96 97.51 33.71 最小值 69.76 65.81 21.47 平均值±
标准差87.17±8.08 81.83±7.70 27.77±3.28 变异系数CV (%) 0.09 0.09 0.12 浅水养殖的栉孔扇贝
C. farreri from the shallow water最大值 74.54 70.13 26.46 最小值 65.86 59.67 19.75 平均值±
标准差69.16±2.22 65.11±2.50 22.959±1.62 变异系数CV (%) 0.03 0.04 0.07 各性状的独立样本T检验见表 3。深水养殖的栉孔扇贝的壳高、壳长和壳宽都极显著大于浅水养殖的栉孔扇贝(P<0.01)。
表 3 传统形态测量学的独立样本T检验Table 3. Independent-samples T test results of traditional morphometry指标Index 独立样本T检验Independent-samples T test 自由度df 显著性P 壳高SH 33.34 2.13×10–13 壳长SL 35.06 2.97×10–13 壳宽SW 42.29 7.17×10–9 注: **表示差异极显著(P<0.01)Note: **P<0.01 is significantly different 2.2 几何形态测量
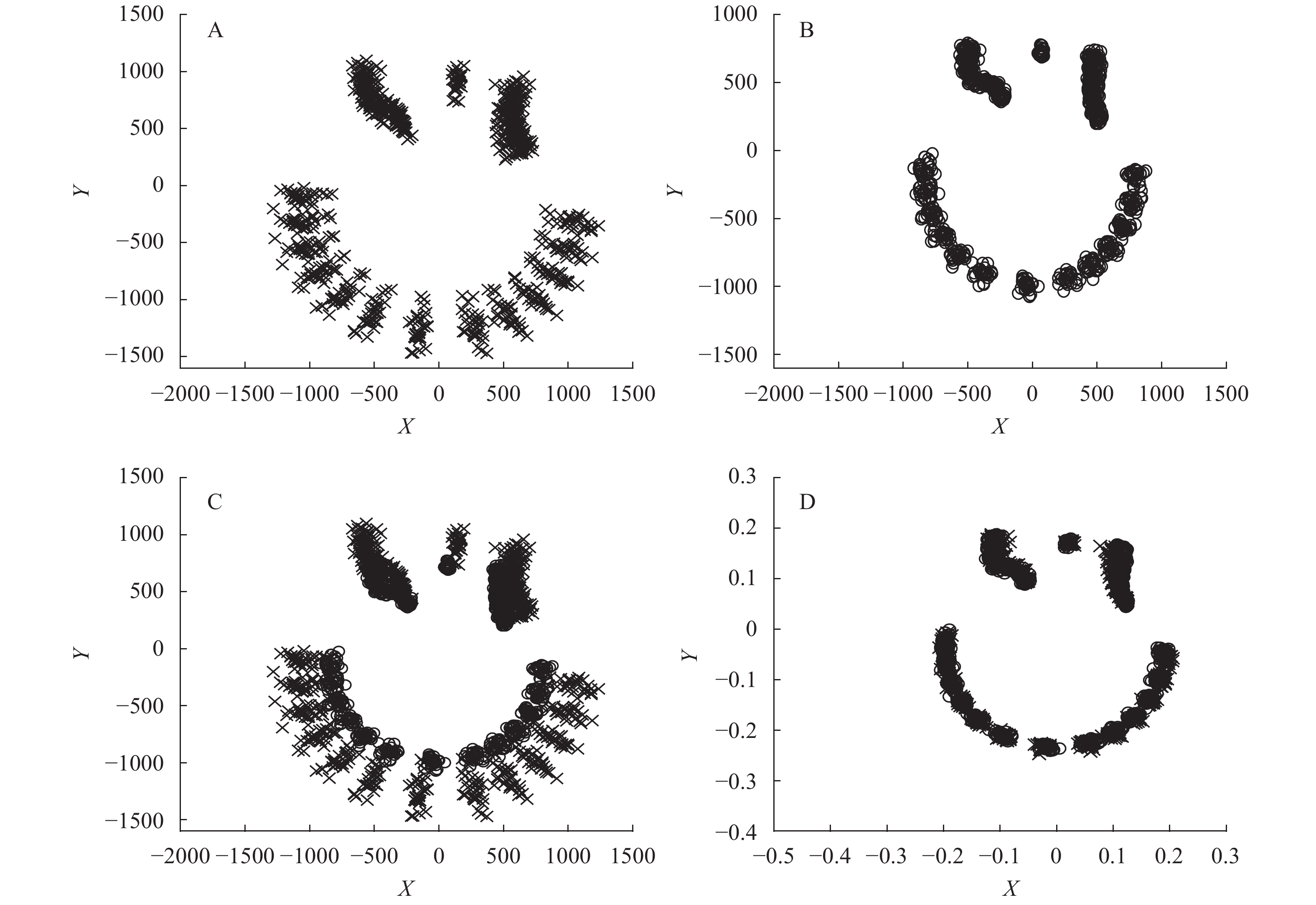
使用PAST软件进行广义普鲁克分析, 在X-Y坐标轴上作2种不同水深栉孔扇贝的GPA叠加图(图 3)。图 3A为深水栉孔扇贝GPA叠加图, 可以看出, 深水栉孔扇贝的壳顶标点较集中, 仅有部分个体的标记点离散于重叠点; 前耳足丝孔内陷程度较一致, 后耳弯曲的倾斜程度不一; 壳缘大小不一, 也有部分个体离散于重叠点。图 3B为浅水栉孔扇贝GPA叠加图, 壳顶标点集中; 前耳集中成一线内折; 后耳线段上粗下细, 说明后耳末端标点较离散; 后耳拐弯处标点较集中; 壳缘标点集中, 大小一致。图 3C为深水栉孔扇贝和浅水栉孔扇贝的GPA叠加图, 可以看出, 浅水栉孔扇贝的壳顶叠加点在深水栉孔扇贝的内侧; 前耳和后耳都向内倾斜和弯曲; 浅水栉孔扇贝的壳缘叠加点都在深水栉孔扇贝壳缘叠加点的内侧。图 3D为去除尺寸大小因素后两种扇贝的GPA叠加图, 可以看出浅水栉孔扇贝和深水栉孔扇贝的壳顶都集中于一点; 前耳弯曲程度一致; 后耳末端标点离散, 但拐弯处标点集中; 壳缘标点集中, 大小一致。
![]() 图 3 不同水深栉孔扇贝的广义普鲁克分析(GPA)叠加图A. 深水栉孔扇贝GPA叠加图; B. 浅水栉孔扇贝GPA叠加图; C. 深水栉孔扇贝和浅水栉孔扇贝的GPA叠加图; D. 去除尺寸大小因素后两种扇贝的GPA叠加图; ×. 深水栉孔扇贝; ○. 浅水栉孔扇贝Figure 3. GPA-landmark plot of C. farreri from different water depthsA. GPA-landmark plot of the scallops from the deep water; B. GPA-landmark plot of the scallops from the shallow water; C. GPA-landmark plot of the scallops from the deep water and the shallow water; D. GPA-landmark plot of the scallops from the deep water and the shallow water after removing the size factor; ×. C. farreri from the deep water; ○. C. farreri from the shallow water
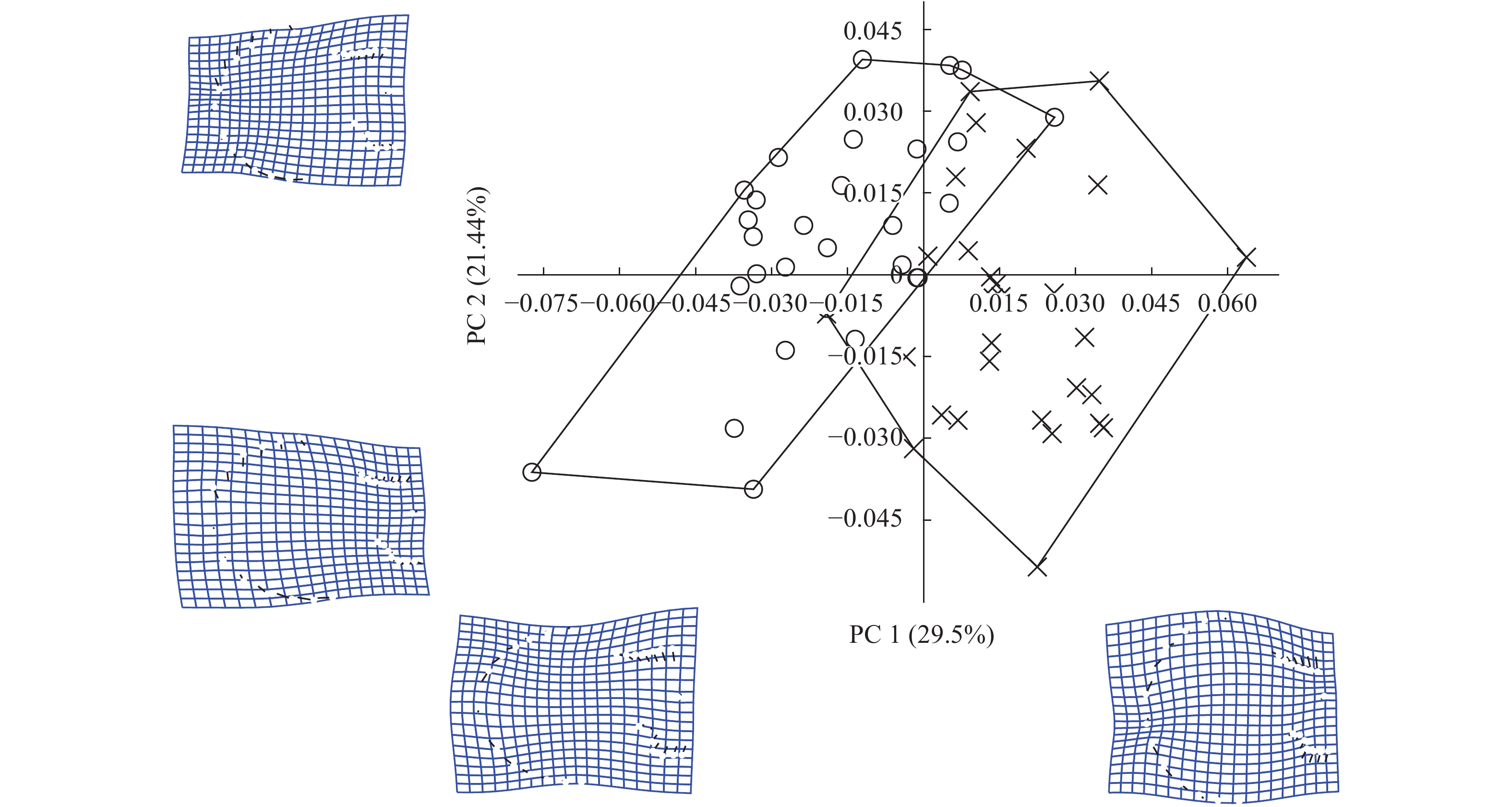
图 3 不同水深栉孔扇贝的广义普鲁克分析(GPA)叠加图A. 深水栉孔扇贝GPA叠加图; B. 浅水栉孔扇贝GPA叠加图; C. 深水栉孔扇贝和浅水栉孔扇贝的GPA叠加图; D. 去除尺寸大小因素后两种扇贝的GPA叠加图; ×. 深水栉孔扇贝; ○. 浅水栉孔扇贝Figure 3. GPA-landmark plot of C. farreri from different water depthsA. GPA-landmark plot of the scallops from the deep water; B. GPA-landmark plot of the scallops from the shallow water; C. GPA-landmark plot of the scallops from the deep water and the shallow water; D. GPA-landmark plot of the scallops from the deep water and the shallow water after removing the size factor; ×. C. farreri from the deep water; ○. C. farreri from the shallow water经GPA变换后, 对样本数据的主成分分析结果中, 前3个主成分(Principal Component, PC)的累计贡献率达到了62.01%, 能够说明2种不同水深栉孔扇贝的主要形态差异, 其中PC1的贡献率为29.52%, PC2的贡献率为21.44%, 以贡献值最大的两个主成分为坐标作二维分布散点图(图 4)。PC1的主要变异位点为2、7—18、21—26, 即扇贝的前后耳和扇形顶部边缘; PC2的主要变异位点为4、6、8—11、19—21、24—28, 即扇贝的后耳和扇形左右部分的下边缘。以扇贝中轴中心点为参照, 当PC1逐渐增大时, 扇贝前后耳呈向外变化、扇形顶部边缘呈向下变化的趋势; 当PC2逐渐增大时, 扇贝后耳呈向内变化、扇形左边缘呈向外变化、扇形右边缘呈向下变化的趋势。图中深水栉孔扇贝主要分布在PC1的正轴和PC2的负轴, 即壳耳呈向外变化、扇形顶部边缘呈向下变化、左边缘呈向内变化、右边缘呈向上变化的趋势; 浅水栉孔扇贝主要分布在PC1的负轴和PC2的正轴, 即壳耳呈向内变化、扇形顶部边缘呈向上变化、左边缘呈向外变化、右边缘呈向下变化的趋势。
![]() 图 4 不同水深栉孔扇贝的主成分分析(PCA)散点图×. 深水栉孔扇贝; ○. 浅水栉孔扇贝Figure 4. Principal components analysis (PCA) diagram of two groups×. C. farreri from the deep water; ○. C. farreri from the shallow water
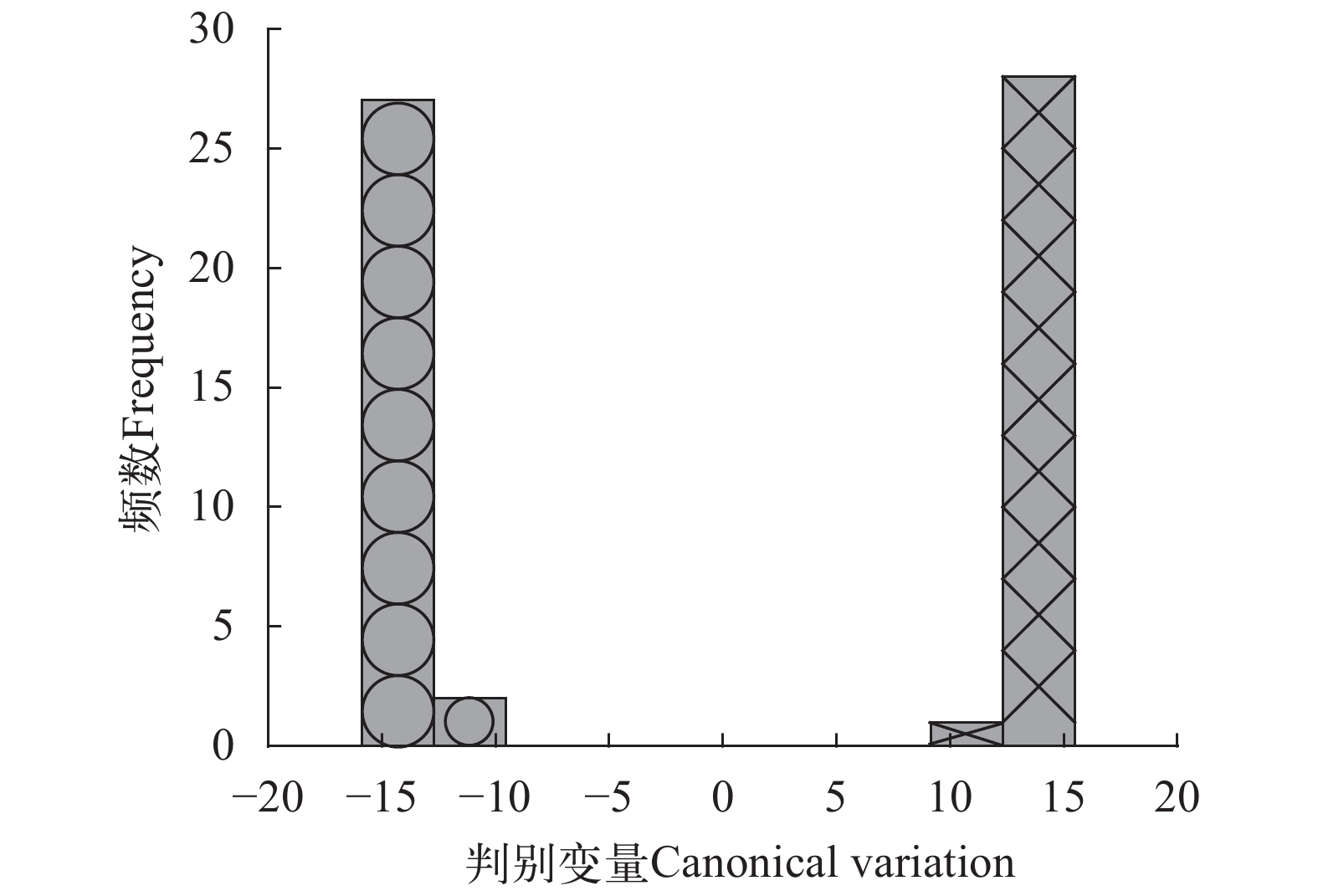
图 4 不同水深栉孔扇贝的主成分分析(PCA)散点图×. 深水栉孔扇贝; ○. 浅水栉孔扇贝Figure 4. Principal components analysis (PCA) diagram of two groups×. C. farreri from the deep water; ○. C. farreri from the shallow water对样本数据进行典型变量分析。先是将数据分类, 从合并的组内协方差矩阵计算马氏距离, 将每个样本分配给马氏距离组平均值最小的组, 得出线性判别分类结果。以判别变量为横轴, 样本频数为纵轴, 作判别直方图(图 5)。图中可见2种不同水深的栉孔扇贝在坐标上分为两簇, 判别率为100%, 说明两者之间通过典型变量分析能够独立分开。
![]() 图 5 不同水深栉孔扇贝的典型变量分析(CVA)直方图×. 深水栉孔扇贝; ○. 浅水栉孔扇贝Figure 5. Canonical variate analysis (CVA) histogram of the two groups×. C. farreri from the deep water; ○. C. farreri from the shallow water
图 5 不同水深栉孔扇贝的典型变量分析(CVA)直方图×. 深水栉孔扇贝; ○. 浅水栉孔扇贝Figure 5. Canonical variate analysis (CVA) histogram of the two groups×. C. farreri from the deep water; ○. C. farreri from the shallow water3. 讨论
在形态测量学中, 传统形态测量学与几何形态测量学相比存在着统计学上的缺陷[34]。传统形态测量学方法的度量值通常与被测物的尺寸大小高度相关[35], 分析结果对所选的特定距离或比率非常敏感, 所以通过传统测量方法很难得到稳健的不含尺寸的形状变量, 而几何形态测量法在分析过程中保留了形状变量[36-38], 能通过坐标变换分析得到非尺寸大小的形状差异。比如Márquez等[32]对特维尔切扇贝Aequipecten tehuelchus的几何形态学研究发现, 该扇贝在从稚贝开始的5年生长过程中, 为适应从附着到游泳生活的习性, 前后耳从对称变为不对称、壳形从细长变为椭圆; 王贺崐元等[39]通过对高原裸裂尻鱼头部轮廓的几何形态测量分析, 得到了其不同亚种和居群间非尺寸大小性状的形态多样性。
本研究一是通过传统形态测量学的线性距离统计学方法, 分析得到深水养殖的栉孔扇贝的壳高(SH)、壳长(SL)和壳宽(SW)极显著大于浅水养殖的栉孔扇贝(P<0.01); 二是通过几何形态测量学的数字标点多变量统计方法, 分析得到深水养殖的栉孔扇贝的整体壳形尺寸大于浅水养殖的栉孔扇贝, 并且发现除去尺寸大小因素后两者的壳耳和扇形边缘存在显著差异。两种分析方法都能得到深水养殖栉孔扇贝壳形尺寸大于浅水养殖的栉孔扇贝的结果, 且几何形态测量学能得到更加具体和详细的形态变化趋势。
王光花等[30]对桑沟湾远岸水域(30 m水深)养殖的栉孔扇贝的数量性状的相关性和通径分析, 在远岸深水区养殖栉孔扇贝有较高的经济效益。对花蛤的深水养殖研究表明, 比起传统的养殖方式, 其成活率和平均产量都显著提升, 投入产出比为1﹕4.5, 增加收益12.6%[40], 由此可见, 深水养殖是贝类养殖增加效益的可靠方式, 而且深水养殖对拓展养殖海域、减轻近岸养殖区域的环境压力、调节和保护海洋养殖品种结构都有着重要意义, 是我国实现海水养殖生态经济协调发展的必然趋势[41]。
形态测量学的发展是计算机技术和数学方法的创新和完善[38]。一方面随着3D技术的兴起, 在能够处理3D图像和三维数据的软件支持下, 对生物体立体形态的分析成为可能, 分析角度更加多元、形态更加细节; 另一方面其分析方法也融合了数学、物理等方面的知识, 熟练掌握此方法需要扎实的自然科学功底和较好的理论基础。在今后的研究中, 掌握相关数理知识, 利用先进的计算机科学技术, 从不同领域、站在不同角度解释和分析生物体或生物体间的形态变化和差异, 也是该学科未来的发展思路[42, 43]。
-
![]()
图 1 栉孔扇贝传统形态测量学指标
1. 深水栉孔扇贝; 2. 浅水栉孔扇贝
Figure 1. Traditional morphometrical measurements of C. farreri
1. The scallops in the deep water; 2. The scallops in the shallow water
![]()
图 2 栉孔扇贝界标点与半界标点的位置
Figure 2. Position of the 8 landmarks and 20 semi-landmarks on C. farreri
![]()
图 3 不同水深栉孔扇贝的广义普鲁克分析(GPA)叠加图
A. 深水栉孔扇贝GPA叠加图; B. 浅水栉孔扇贝GPA叠加图; C. 深水栉孔扇贝和浅水栉孔扇贝的GPA叠加图; D. 去除尺寸大小因素后两种扇贝的GPA叠加图; ×. 深水栉孔扇贝; ○. 浅水栉孔扇贝
Figure 3. GPA-landmark plot of C. farreri from different water depths
A. GPA-landmark plot of the scallops from the deep water; B. GPA-landmark plot of the scallops from the shallow water; C. GPA-landmark plot of the scallops from the deep water and the shallow water; D. GPA-landmark plot of the scallops from the deep water and the shallow water after removing the size factor; ×. C. farreri from the deep water; ○. C. farreri from the shallow water
![]()
图 4 不同水深栉孔扇贝的主成分分析(PCA)散点图
×. 深水栉孔扇贝; ○. 浅水栉孔扇贝
Figure 4. Principal components analysis (PCA) diagram of two groups
×. C. farreri from the deep water; ○. C. farreri from the shallow water
![]()
图 5 不同水深栉孔扇贝的典型变量分析(CVA)直方图
×. 深水栉孔扇贝; ○. 浅水栉孔扇贝
Figure 5. Canonical variate analysis (CVA) histogram of the two groups
×. C. farreri from the deep water; ○. C. farreri from the shallow water
表 1 栉孔扇贝界标点与半界标点的位置
Table 1 The position of landmarks and semi-landmarks on C. farreri
标点Landmark and Semi-landmark 对应部位Position 1 壳顶 2 前耳前端 3 足丝孔 4 贝壳右边缘最宽处 5 壳底 6 贝壳左边缘最宽处 7 后耳拐弯处 8 后耳末端 9—13 沿着后耳轮廓线的半界标点 14—18 沿着前耳轮廓线的半界标点 19—23 沿着贝壳右边缘轮廓线的半界标点 24—28 沿着贝壳左边缘轮廓线的半界标点  下载: 导出CSV
下载: 导出CSV
表 2 传统形态测量学的基本统计量
Table 2 Basic statistics of traditional morphometry
种类Specimen 统计量
Statistic壳高SH (mm) 壳长SL (mm) 壳宽SW (mm) 深水养殖的栉孔扇贝
C. farreri from the deep water最大值 102.96 97.51 33.71 最小值 69.76 65.81 21.47 平均值±
标准差87.17±8.08 81.83±7.70 27.77±3.28 变异系数CV (%) 0.09 0.09 0.12 浅水养殖的栉孔扇贝
C. farreri from the shallow water最大值 74.54 70.13 26.46 最小值 65.86 59.67 19.75 平均值±
标准差69.16±2.22 65.11±2.50 22.959±1.62 变异系数CV (%) 0.03 0.04 0.07
下载: 导出CSV
表 3 传统形态测量学的独立样本T检验
Table 3 Independent-samples T test results of traditional morphometry
指标Index 独立样本T检验Independent-samples T test 自由度df 显著性P 壳高SH 33.34 2.13×10–13 壳长SL 35.06 2.97×10–13 壳宽SW 42.29 7.17×10–9 注: **表示差异极显著(P<0.01)Note: **P<0.01 is significantly different
下载: 导出CSV
-
[1] D’Arcy W T. On Growth and Form [M]. Cambridge: Cambridge University Press, 1917: 719.
[2] 王旭, 徐衡, 邹莉, 等. 3个单齿螺地理群体形态差异分析 [J]. 南方水产科学, 2013, 9(4): 22-27. doi: 10.3969/j.issn.2095-0780.2013.04.004 Wang X, Xu H, Zou L, et al. Morphological variation analysis of three different geographic populations of Monodonta labio [J]. South China Fisheries Science, 2013, 9(4): 22-27. [ doi: 10.3969/j.issn.2095-0780.2013.04.004
[3] 牛泓博, 聂鸿涛, 赵力强, 等. 辽宁沿海菲律宾蛤仔不同地理群体形态差异研究 [J]. 海洋科学, 2015, 39(11): 54-60. doi: 10.11759/hykx20150731002 Niu H B, Nie H T, Zhao L Q, et al. Morphological variation analysis of the Manila clam (Ruditapes philippinarum) populations along the coast of Liaoning Province [J]. Marine Sciences, 2015, 39(11): 54-60. [ doi: 10.11759/hykx20150731002
[4] MacLeod N. Geometric morphometrics and geological shape-classification systems [J]. Earth-Science Reviews, 2002, 59(1-4): 27-47. doi: 10.1016/S0012-8252(02)00068-5
[5] 来益同, 沈华, 张奕祥, 等. 2个纯合体色品系大瓶螺的几何形态测量学研究 [J]. 水生态学杂志, 2016, 37(2): 88-95. Lai Y T, Shen H, Zhang Y X, et al. Geometric morphometric study of two homozygous color strains of the golden apple snail Pomacea canaliculate (Caenogastropoda: Ampullariidae) [J]. Journal of Hydroecology, 2016, 37(2): 88-95. [
[6] 陈楠桦, 梁仁杰, 白义, 等. 基于几何形态测量学的四种滨螺形态差异与系统发生关系研究 [J]. 海洋与湖沼, 2018, 49(6): 239-248. Chen N H, Liang R J, Bai Y, et al. A geometric-morphometry-based study on phylogeny of four Littorina (Gastropoda: Littorinidae) species [J]. Oceanologia et Limnologia Sinica, 2018, 49(6): 239-248. [
[7] 李成林, 宋爱环, 胡炜, 等. 山东省扇贝养殖产业现状分析与发展对策 [J]. 海洋科学, 2011, 35(3): 92-98. Li C L, Song A H, Hu W, et al. Status analyzing and developing counter-measure of cultured scallop industry in Shandong Province [J]. Marine Sciences, 2011, 35(3): 92-98. [
[8] 张福绥, 杨红生. 山东沿岸夏季栉孔扇贝大规模死亡原因分析 [J]. 海洋科学, 1999, 23(1): 44-47. doi: 10.3969/j.issn.1000-3096.1999.01.018 Zhang F S, Yang H S. Analysis of the causes of mass mortality of farming Chlamys farreri in summer in coastal areas of Shandong, China [J]. Marine Sciences, 1999, 23(1): 44-47. [ doi: 10.3969/j.issn.1000-3096.1999.01.018
[9] 蒋增杰. 桑沟湾栉孔扇贝大规模死亡原因分析 [J]. 齐鲁渔业, 2004(8): 10-11. Jiang Z J. Analysis of the causes of mass mortality of Chlamys farreri in Sanggou Bay [J]. Shandong Fisheries, 2004(8): 10-11. [
[10] 杨世尧. 昌黎扇贝养殖走向深海 [J]. 农产品市场周刊, 2016(45): 11. doi: 10.3969/j.issn.1009-8070.2016.45.007 Yang S Y. Moving towards deep-sea scallop cultured in Changli [J]. Agricultural Products Market Weekly, 2016(45): 11. [ doi: 10.3969/j.issn.1009-8070.2016.45.007
[11] 操戈, 邓卫哲, 徐承旭. 海南大力发展深海网箱养殖 [J]. 水产科技情报, 2017(2): 60. Cao G, Deng W Z, Xu C X. Hainan vigorously develops deep-sea cage culture [J]. Fisheries Science & Technology Information, 2017(2): 60. [
[12] 侯海燕, 鞠晓晖, 陈雨生. 国外深海网箱养殖业发展动态及其对中国的启示 [J]. 世界农业, 2017(5): 162-166. Hou H Y, Ju X H, Chen Y S. Development trends of deep-sea cage farming industry in foreign countries and its enlightenment to China [J]. World Agriculture, 2017(5): 162-166. [
[13] Stanley S M. Relation of Shell Form to Life Habits of the Bivalvia (Mollusca) [M]. McLean: Geological Society of America, 1970: 1-296
[14] Gould S J. Muscular mechanics and the ontogeny of swimming in scallops [J]. Palaeontology, 1971, 14(Part 1): 61-94.
[15] Ackerly S C. The structure of ontogenetic variation in the shell of Pecten [J]. Palaeontology, 1992(34): 847-867.
[16] Manuel J L, Dadswell M J. Swimming of juvenile sea scallops, Placopecten magellanicus (Gmelin): a minimum size for effective swimming [J]. Journal of Experimental Marine Biology and Ecology, 1993, 174(2): 137-175. doi: 10.1016/0022-0981(93)90015-G
[17] Gaspar M B, Santos M N, Vasconcelos P, et al. Shell morphometric relationships of the most common bivalve species (Mollusca: Bivalvia) of the Algarve coast (southern Portugal) [J]. Hydrobiologia, 2002, 477(1-3): 73-80.
[18] Ubukata T. A morphometric study on morphological plasticity of shell form in crevice-dwelling Pterioida (Bivalvia) [J]. Biological Journal of the Linnean Society, 2003, 79(2): 285-297. doi: 10.1046/j.1095-8312.2003.00144.x
[19] Márquez F, Robledo J, Peñaloza G E, et al. Use of different geometric morphometrics tools for the discrimination of phenotypic stocks of the striped clam Ameghinomya antiqua (Veneridae) in north Patagonia, Argentina [J]. Fisheries Research, 2010, 101(1-2): 127-131. doi: 10.1016/j.fishres.2009.09.018
[20] Márquez F, González-José R, Bigatti G. Combined methods to detect pollution effects on shell shape and structure in Neogastropods [J]. Ecological Indicators, 2011, 11(2): 248-254. doi: 10.1016/j.ecolind.2010.05.001
[21] 刘小林, 常亚青, 相建海, 等. 栉孔扇贝壳尺寸性状对活体重的影响效果分析 [J]. 海洋与湖沼, 2002, 33(6): 673-678. doi: 10.3321/j.issn:0029-814X.2002.06.014 Liu X L, Chang Y Q, Xiang J H, et al. Analysis of effects of shell size characters on live weight in Chinese scallop Chlamys farreri [J]. Oceanologia Et Limnologia Sinica, 2002, 33(6): 673-678. [ doi: 10.3321/j.issn:0029-814X.2002.06.014
[22] 钱旭阳, 张永普. 泥蚶数量性状的相关分析 [J]. 温州师范学院学报(自然科学版), 2004, 25(2): 84-86. Qian X Y, Zhang Y P. The quantitative characters correlated response of Tegillarca granosa [J]. Journal of Wenzhou Teachers College, 2004, 25(2): 84-86. [
[23] 张存善, 常亚青, 曹学彬, 等. 虾夷扇贝体形性状对软体重和闭壳肌重的影响效果分析 [J]. 水产学报, 2009, 33(1): 87-94. Zhang C S, Chang Y Q, Cao X B, et al. Analysis of effect of body shape traits on soft-tissue weight and adductor weight in Patinopecten yessoensis Jay [J]. Journal of Fisheries of China, 2009, 33(1): 87-94. [
[24] 刘志刚, 章启忠, 王辉. 华贵栉孔扇贝主要经济性状对闭壳肌重的影响效果分析 [J]. 热带海洋学报, 2009, 28(1): 61-66. doi: 10.3969/j.issn.1009-5470.2009.01.010 Liu Z G, Zhang Q Z, Wang H. Effects of main economic traits on adductor weight of Chlamys nobilis (Reeve) [J]. Journal of Tropical Oceanography, 2009, 28(1): 61-66. [ doi: 10.3969/j.issn.1009-5470.2009.01.010
[25] 王庆恒, 邓岳文, 杜晓东, 等. 翡翠贻贝形态性状对软体部质量的影响 [J]. 广东海洋大学学报, 2009, 29(4): 1-4. doi: 10.3969/j.issn.1673-9159.2009.04.001 Wang Q H, Deng Y W, Du X D, et al. Effect of morphometric traits on edible part weight of Perna viridis [J]. Journal of Guangdong Ocean University, 2009, 29(4): 1-4. [ doi: 10.3969/j.issn.1673-9159.2009.04.001
[26] 孙泽伟, 郑怀平, 杨彦鸿, 等. 近江牡蛎养殖群体数量性状间的相关及通径分析 [J]. 中国农学通报, 2010, 26(6): 332-336. Sun Z W, Zheng H P, Yang Y H, et al. Correlation and path analysis to quantitative traits for a cultured population of Jinjiang oyster Crassostrea hongkongensis [J]. Chinese Agricultural Science Bulletin, 2010, 26(6): 332-336. [
[27] 杜美荣, 王彬, 张继红, 等. 一龄栉孔扇贝壳长与壳高对湿重的相关性和通径分析 [J]. 中国农学通报, 2012, 28(20): 136-139. doi: 10.11924/j.issn.1000-6850.2012-0895 Du M R, Wang B, Zhang J H, et al. Correlation and path analysis on shell length and shell heightto wet weight of Chlamys farreri at one-year old [J]. Chinese Agricultural Science Bulletin, 2012, 28(20): 136-139. [ doi: 10.11924/j.issn.1000-6850.2012-0895
[28] 于宗赫. 海州湾深水区域两种养殖方式栉孔扇贝生长与免疫特征的比较 [C]. 中国动物学会贝类学分会、中国海洋湖沼学会贝类学分会、中国动物学会、中国海洋湖沼学会贝类学会分会第十四次学会研讨会论文摘要汇编. 中国动物学会贝类学分会、中国海洋湖沼学会贝类学分会: 中国海洋湖沼学会, 2009: 1. Yu Z H. Comparison of growth and immune characteristics of chlamys farreri by two culture methods in deep-water area of haizhou bay [C]. China Zoological Society Molocologia Branch, Chinese Society for Oceanology and Limnology Molocologia Branch. Compilation of Abstracts of the Fourteenth Society Symposium of China Zoological Society & Chinese Society for Oceanology and Limnology Molocologia Branch. China Zoological Society Molocologia Branch, Chinese Society for Oceanology and Limnology Molocologia Branch: Chinese Society for Oceanology and Limnology, 2009: 1
[29] 张继红, 方建光, 唐启升, 等. 桑沟湾不同区域养殖栉孔扇贝的固碳速率 [J]. 渔业科学进展, 2013, 34(1): 12-16. doi: 10.3969/j.issn.1000-7075.2013.01.002 Zhang J H, Fang J G, Tang Q S, et al. Carbon sequestration rate of the scallop Chlamys farreri cultivated in different areas of Sanggou Bay [J]. Progress in Fishery Sciences, 2013, 34(1): 12-16. [ doi: 10.3969/j.issn.1000-7075.2013.01.002
[30] 王光花, 蒋增杰, 方建光. 浅海远岸水域筏养栉孔扇贝数量性状的相关性及通径分析 [J]. 现代农业科技, 2011(19): 327-328. doi: 10.3969/j.issn.1007-5739.2011.19.215 Wang G H, Jiang Z J, Fang J G. Correlation and path analysis on quantitative traits of Chlamys farreri in offshore area [J]. Modern Agricultural Science and Technology, 2011(19): 327-328. [ doi: 10.3969/j.issn.1007-5739.2011.19.215
[31] Mullin S K, Taylor P J. The effects of parallax on geometric morphometric data [J]. Computers in Biology and Medicine, 2002, 32(6): 455-464. doi: 10.1016/S0010-4825(02)00037-9
[32] Márquez F, Amoroso R, Sainz M F G, et al. Shell morphology changes in the scallop Aequipecten tehuelchus during its life span: a geometric morphometric approach [J]. Aquatic Biology, 2010, 11(2): 149-155. doi: 10.3354/ab00301
[33] Hammer O, Harper D A T, Ryan P D. PAST: paleontological statistics software package for education and data analysis [J]. Palaeontologia Electronica, 2001, 4(1): 9.
[34] Sokal R R, Rohlf F J. Biometry [M]. New York: WH Freeman and Company, 1995: 578.
[35] Bookstein F L. Morphometric Tools for Landmark Data: Geometry and Biology [M]. New York: Cambridge University Press, 1991: 435.
[36] Rohlf F J, Marcus L F. A revolution morphometrics [J]. Trends in Ecology & Evolution, 1993, 8(4): 129-132.
[37] Adams D C, Rohlf F J, Slice D E. Geometric morphometrics: ten years of progress following the ‘revolution’ [J]. Italian Journal of Zoology, 2004, 71(1): 5-16. doi: 10.1080/11250000409356545
[38] Mitteroecker P, Gunz P. Advances in geometric morphometrics [J]. Evolutionary Biology, 2009, 36(2): 235-247. doi: 10.1007/s11692-009-9055-x
[39] 王贺崐元, 汤永涛, 李国刚, 等. 高原裸裂尻鱼头部轮廓形状及其变异的几何形态测量分析 [J]. 水生生物学报, 2017, 41(1): 182-193. doi: 10.7541/2017.23 Wang H K Y, Yang Y T, Li G G, et al. Geometric morphometrics of the cephalic contour and its morphological variations among Schizopygopsis stoliczkai (Teleostei: Cyprinidae) [J]. Acta Hydrobiologica Sinica, 2017, 41(1): 182-193. [ doi: 10.7541/2017.23
[40] 乔曦. 深水养殖花蛤研究与应用 [J]. 水产养殖, 2008, 29(6): 21-22. doi: 10.3969/j.issn.1004-2091.2008.06.007 Qiao X. Research and application of deep-water aquaculture clam [J]. Journal of Aquaculture, 2008, 29(6): 21-22. [ doi: 10.3969/j.issn.1004-2091.2008.06.007
[41] 余粮红, 郑珊, 高强. 国外海水养殖生态经济系统协调模式及其对中国的启示 [J]. 世界农业, 2018(7): 112-120. Yu L H, Zheng S, Gao Q. Coordination models of maricultural eco-economic systems in foreign countries and their implications for China [J]. World Agriculture, 2018(7): 112-120. [
[42] Richtsmeier J T, Burke Deleon V, Lele S R. The promise of geometric morphometrics [J]. American Journal of Physical Anthropology, 2002, 119(S35): 63-91. doi: 10.1002/ajpa.10174
[43] Lawing A M, Polly P D. Geometric morphometrics: recent applications to the study of evolution and development [J]. Journal of Zoology, 2010, 280(1): 1-7. doi: 10.1111/j.1469-7998.2009.00620.x
-
期刊类型引用(10)
1. 金武,马学艳,彭刚,陈琬雯,闻海波. 梨形环棱螺11个地理种群的几何形态学分析. 中国农学通报. 2024(02): 159-164 .  百度学术
百度学术
2. Ziniu ZHANG,Zhenyan WANG,Guihua LI,Meihan ZHAO,Wenjian LI. The suitability assessment on site selection for bottom-seeding scallop culture based on analytic hierarchy process. Journal of Oceanology and Limnology. 2024(02): 647-663 . 必应学术
3. 周用武,马骏,詹庆斌. 几何形态测量学在三种鹦鹉蛋识别中的应用与展望. 山西师范大学学报(自然科学版). 2024(01): 76-82 . 百度学术
4. 巩梦,骆劲舟,代乔坤,刘紫薇,张崧,白琳,华洪. 华北陆块西南缘寒武系辛集组Stenotheca型和Pelagiella型壳体化石几何形态测量学初探. 微体古生物学报. 2024(01): 1-13 . 百度学术
5. 黄会,刘小静,刘慧慧,张健,徐英江,李佳蔚,王共明,姜立生. 栉孔扇贝蒸煮液营养成分分析与安全性评价. 中国食品添加剂. 2023(03): 250-256 . 百度学术
6. 赵春暖,于涛,郑言鑫,李波,王喜文,蔡忠强,王晓梅,任利群,徐帅,吴玉萍,吴楠,张明燡. 不同壳色和性别栉孔扇贝群体的性状相关性及通径分析. 中国水产科学. 2023(03): 268-283 . 百度学术
7. 于道德,刘凯凯,张少春,邱兆星. 中国栉孔扇贝养殖和育种的回顾与展望. 海洋科学. 2023(08): 112-119 . 百度学术
8. Kaiyu Yang,Jianghao Wu,Xinman Li,Xinbo Pang,Yangchen Yuan,Guohui Qi,Minsheng Yang. Intraspecific leaf morphological variation in Quercus dentata Thunb.: a comparison of traditional and geometric morphometric methods, a pilot study. Journal of Forestry Research. 2022(06): 1751-1764 . 必应学术
9. 雷菁,万智巍,鞠民. 基于形态计量学和CCA分析的杂粮淀粉粒识别与鉴定. 食品工业科技. 2021(14): 289-295 . 百度学术
10. 刘澜鼎,王伟,梅耀杰,刘合叶,崔龙波. 烟台四十里湾海域养马岛贝类养殖区水质监测与评价. 渔业研究. 2021(04): 394-402 . 百度学术
其他类型引用(5)


计量
- 文章访问数: 3579
- HTML全文浏览量: 682
- PDF下载量: 73
- 被引次数: 15