
EFFECTS OF DIFFERENT PROTEIN SOURCES ON GROWTH PERFORMANCE AND METABOLISM-RELATED GENE EXPRESSION IN JUVENILE DARKBARBEL CATFISH (PELTEOBAGRUS VACHELLI)
-
摘要: 以初始体重为(2.90±0.01) g的瓦氏黄颡鱼(Pelteobagrus vachelli)为研究对象, 探究不同蛋白源对瓦氏黄颡鱼生长、体组成、前肠PEPT1和肝脏TOR基因表达的影响。以鱼粉(FM)、大豆浓缩蛋白(SPC)、水解鱼蛋白(FH)和晶体氨基酸混合物(CAA)为主要蛋白源, 配制4种等氮(粗蛋白含量为39.0%)等脂(粗脂肪含量为9.0%)的实验饲料, 投喂实验鱼66d。结果显示, 不同蛋白源虽然对瓦氏黄颡鱼存活率(SR)无显著影响(P>0.05), 但显著影响了瓦氏黄颡鱼的特定生长率(SGR)。其中, SPC组瓦氏黄颡鱼SGR低于FM组, 却显著高于FH组和CAA组(P<0.05)。虽然SPC组瓦氏黄颡鱼粗蛋白含量低于FM组, 但其鱼体粗脂肪含量显著高于FM组(P<0.05)。不同蛋白源显著影响了瓦氏黄颡鱼PEPT1基因表达(P<0.05), 在35d时, SPC组PEPT1表达量虽然低于FM组, 但显著高于FH和CAA组(P<0.05);在66d时, SPC组瓦氏黄颡鱼前肠PEPT1表达量显著高于FM组(P<0.05)。不同蛋白源饲料对鱼体肝脏TOR基因表达无显著影响(P>0.05)。以上结果表明, SPC组瓦氏黄颡鱼生长性能虽然低于FM组, 但显著高于FH组和CAA组, 这可能是通过上调前肠PEPT1表达来实现的。Abstract: A 66-day feeding trial was conducted to investigate the effects of different protein sources on growth, body composition and PEPT1 gene expression in the foreintestine and TOR gene expression in the liver of juvenile darkbarbel catfish [initial weight (2.90 ± 0.01) g]. Four isoproteic (39.0% crude protein) and isolipidic (9.0% crude lipid) diets were formulated with fish meal (FM), soy protein concentrate (SPC), fish hydrolysate (FH) and crystal amino acids (CAA) as the main protein source. No significant difference was observed in the survival rate among dietary treatments (P>0.05). Dietary protein had a significant effect on SGR. SPC group had lower SGR than that of the FM groups, while SPC group had higher SGR than FH and CAA group (P<0.05). The crude protein of SPC group was only lower than the FM group, while the crude lipid of SPC group was higher than the FM group (P<0.05). Dietary protein had a significant effect on PEPT1 mRNA expression. The PEPT1 mRNA expression in SPC group was significantly lower than the FM group, but significantly higher than the FH and CAA group at day 35. However, the PEPT1 mRNA expression of SPC group was significantly higher than the FM group at day 66 (P<0.05). Dietary protein source had no effect on TOR gene expression. These results suggested that juvenile darkbarbel catfish of SPC group had better growth performance than FH and CAA group, which might be attributed to the higher foregut PEPT1 mRNA expression.
-
Keywords:
- Darkbarbel catfish /
- Soy protein concentrate /
- Growth /
- Body composition /
- Gene expression
-
随着集约化养殖的不断发展, 寻找优质蛋白源以替代鱼粉(Fish meal, FM)、节约成本, 成为水产动物营养研究的热点和难点[1—5]。研究发现, 不同蛋白源具有不同的特点, 大豆浓缩蛋白(Soy protein concentrate, SPC)由于蛋白质含量高(65%—70%的粗蛋白), 抗营养因子和纤维含量较低, 同时还具有利用率高、氨基酸平衡性好、价格低廉等优点, 成为极具潜力的鱼粉替代源[6, 7]。在线鳢(Channa striata)[8]和星斑川鲽(Platichthys stellatus Pallas)[9]中研究发现, 用SPC替代部分鱼粉可显著提高鱼体生长率和饲料转化效率。水解鱼肉蛋白(Fish hydrolysate, FH)是一种富含各种肽段、诱食好且促进鱼类生长明显的蛋白源[10, 11]。在大黄鱼(Larimichthys crocea)的养殖中发现, 用过滤后的水解蛋白替代40%的鱼粉, 相比于全鱼粉组, 不影响大黄鱼的存活率和特定生长率[12]。晶体氨基酸(Crystal amino acids, CAA)可被鱼体直接吸收和利用, 因此晶体氨基酸对水产养殖动物来说也是极具价值的鱼粉替代源[13—15]。然而, 这几种蛋白源在同一鱼类中的替代效果尚不明确, 有待进一步研究。
日粮蛋白和内源性蛋白经胰蛋白酶、小肠刷状缘蛋白酶和肽酶等消化后形成小肽和游离氨基酸, 经小肠吸收后进入血液循环, 被机体各组织器官吸收利用[16]。消化后的氨基酸和小肽主要由氨基酸转运载体和小肽转运载体(PEPT1)进行吸收[17]。机体吸收的小肽和氨基酸一方面可用于分解供能, 另一方面可用来重新合成蛋白质, 蛋白质在机体的沉积是由蛋白质的分解和合成共同决定的[18, 19], 其中雷帕霉素靶蛋白(TOR)信号通路对于蛋白质的合成起着至关重要的作用。哺乳动物中关于不同蛋白源调控蛋白质代谢的研究已较为系统[20, 21], 然而鱼类中关于不同蛋白源对PEPT1和TOR基因表达的调控及鱼体生长的影响还有待进一步研究。
瓦氏黄颡鱼(Pelteobagrus vachelli), 鲶形目、鲿科、黄颡鱼属, 温水性鱼类, 我国重要的淡水经济养殖种类之一。近年来, 随着瓦氏黄颡鱼养殖规模的扩大, 对鱼粉的需求量也随之增加, 饲料成本逐渐加大, 因此亟待寻找合适的蛋白源替代鱼粉以促进瓦氏黄颡鱼养殖产业可持续发展[22]。本研究旨在比较不同蛋白源对黄颡鱼生长、体组成和蛋白质代谢基因PEPT1和TOR表达的影响, 以探讨不同蛋白源对黄颡鱼生长性状的影响机理, 为黄颡鱼饲料中鱼粉替代提供理论依据和指导。
1. 材料与方法
1.1 饲料配方和饲料制作
分别以鱼粉(FM)、大豆浓缩蛋白(SPC)、水解鱼蛋白(FH)和晶体氨基酸混合物(CAA)为主要蛋白源, 鱼油、豆油和大豆卵磷脂为主要脂肪源, 小麦粉为糖源制作4种等氮(粗蛋白含量为39.0%)等脂(粗脂肪含量为9.0%)的实验饲料(表 1)。FH购自上海海清饲料公司, 其他饲料原料购自青岛七好生物科技有限公司。
表 1 实验饲料配方和化学成分(%干重)Table 1. Formulation and chemical composition of the experimental diets (% dry matter)原料Ingredient 饲料编号Diet number Diet 1 (FM) Diet 2 (SPC) Diet 3 (FH) Diet 4 (CAA) 鱼粉Fish meal1 48.65 0.00 0.00 0.00 大豆浓缩蛋白SPC1 0.00 48.32 0.00 0.00 晶体氨基酸CAA2 0.00 0.00 0.00 32.50 水解鱼肉蛋白FH1 0.00 0.00 33.69 0.00 小麦粉Wheat gluten1 25.00 25.00 25.00 25.00 麦麸Wheat bran1 13.00 13.00 13.00 13.00 鱼油Fish oil 0.27 4.50 4.41 4.50 豆油Soybean oil 1.81 1.17 1.81 1.81 卵磷脂Lecithin 2.00 2.00 2.00 2.00 矿物质预混料MP3 2.00 2.00 2.00 2.00 维生素预混料VP4 2.00 2.00 2.00 2.00 诱食剂Attractant5 0.50 0.50 0.50 0.50 抗氧化剂Antioxidant 0.05 0.05 0.05 0.05 微晶纤维素MC 4.72 1.46 15.54 16.64 成分分析Proximate composition (% dry matter)5 粗蛋白Crude protein (%) 39.20 39.13 39.30 39.17 粗脂肪Crude lipid (%) 8.60 8.97 9.03 8.79 灰分Ash (%) 6.35 7.33 9.11 10.21 注: 1鱼粉: 粗蛋白占干重66.80%, 粗脂肪占干重8.70%; 大豆浓缩蛋白: 粗蛋白占干重67.26%, 粗脂肪占干重1.31%; 水解鱼肉蛋白: 粗蛋白占干重84.99%, 粗脂肪占干重3.19%; 小麦粉: 粗蛋白占干重16.10%, 粗脂肪占干重1.36%; 麦麸: 粗蛋白占干重19.04%, 粗脂肪占干重2.73%; 2晶体氨基酸(g/kg diet): 精氨酸, 3.42 g; 组氨酸, 0.26 g; 异亮氨酸, 2.52 g; 亮氨酸, 4.36 g; 赖氨酸, 4.26 g; 蛋氨酸, 1.57 g; 苯丙氨酸, 2.29 g; 苏氨酸, 2.41 g; 缬氨酸, 3.06 g; 丙氨酸, 3.67 g; 天冬氨酸, 5.65 g; 胱氨酸, 0.67 g; 谷氨酸, 8.48 g; 甘氨酸, 3.36 g; 丝氨酸, 2.41 g; 酪氨酸, 1.46 g; 脯氨酸, 2.11 g; 3矿物质预混料(mg or g/kg diet): 氟化钠, 2 mg; 碘化钾, 0.8 mg; 氯化钴(1%), 50 mg; 硫酸铜, 10 mg; 硫酸铁, 80 mg; 硫酸锌, 50 mg; 硫酸锰, 60 mg; 硫酸镁, 1200 mg; 磷酸二氢钙, 3000 mg; 氯化钠, 100 mg; 沸石粉, 15.448 g; 4维生素预混料(mg or g/kg diet): 硫胺素, 25 mg; 维生素B2, 45 mg; 盐酸吡哆醇, 20 mg; 维生素B12, 0.1 mg; 维生素K3, 10 mg; 肌醇, 800 mg; 泛酸, 60 mg; 烟酸, 200 mg; 叶酸, 20 mg; 生物素, 1.20 mg; 维生素A, 32 mg; 维生素D, 5 mg; 维生素E, 120 mg; 维生素C, 2000 mg; 胆碱, 2000 mg; 乙氧基喹啉, 150 mg; 微晶纤维素, 14.52 g; 5诱食剂: 甘氨酸和甜菜碱Note: 1Fish meal: crude protein 66.80% dry matter, crude lipid 8.70% dry matter; SPC: crude protein 67.26% dry matter, crude lipid 1.31% dry matter; Fish hydrolysate: crude protein 84.99% dry matter, crude lipid 3.19% dry matter; Wheat gluten: crude protein 16.10% dry matter, crude lipid 1.36% dry matter; Wheat bran: crude protein 19.04% dry matter, crude lipid 2.73% dry matter; 2Crystalline amino acids (g/kg diet): Arginine, 3.42 g; Histidine, 0.26 g; Isoleucine, 2.52 g; Leucine, 4.36 g; Lysine, 4.26 g; Methionine, 1.57 g; Phenylalanine, 2.29 g; Threonine, 2.41 g; Valine, 3.06 g; Alanine, 3.67 g; Aspartic acid, 5.65 g; Cystine, 0.67 g; Glutamic acid, 8.48 g; Glycine, 3.36 g; Serine, 2.41 g; Tyrosine, 1.46 g; Proline, 2.11 g; 3Mineral premix (mg or g/kg diet): NaF, 2 mg; KI, 0.8 mg; CoCl2·6H2O(1%), 50 mg; CuSO4·5H2O, 10 mg; FeSO4·H2O, 80 mg; ZnSO4·H2O, 50 mg; MnSO4·H2O, 60 mg; MgSO4·7H2O, 1200 mg; Ca(H2PO4)2·H2O, 3000 mg; NaCl, 100 mg; Zoelite, 15.448 g; 4Vitamin premix (mg or g/kg diet): thiamin, 25 mg; riboflavin, 45 mg; pyridoxine-HCl, 20 mg; vitamin B12, 0.1 mg; vitamin K3, 10 mg; inositol, 800 mg; pantothenic acid, 60 mg; niacin acid, 200 mg; folic acid, 20 mg; biotin, 1.20 mg; retinol acetate, 32 mg; cholecalciferol, 5 mg; alpha-tocopherol, 120 mg; ascorbic acid, 2000 mg; choline chloride, 2000 mg; ethoxyquin, 150 mg; microcrystalline cellulose, 14.52 g; 5Attractant: glycine and betaine 饲料制作前, 将饲料原料进行粉碎并过246目筛。在饲料制作过程中, 饲料原料根据配方按逐级放大原理混合均匀, 之后加入鱼油、豆油和卵磷脂, 手工将油脂颗粒搓匀, 然后加入适量水, 用全自动渔用饵料机(F-26(Ⅱ), 华南理工大学)进行制粒, 制成直径为1.5 mm×3.0 mm饲料颗粒, 以45℃烘至水分含量小于10%, 用双层塑料袋扎口保存, 置−20℃冰箱中备用。
1.2 养殖实验和样品采集
瓦氏黄颡鱼购于四川眉山育苗场, 实验前, 实验鱼在广州市农业科学院白云基地循环水系统中暂养2周, 以FM组饲料进行饱食投喂, 使实验鱼逐渐适应人工饲料和养殖环境。在暂养结束后, 黄颡鱼饥饿24h, 将初重(2.90±0.01) g的瓦氏黄颡鱼随机分成4组, 每组3个重复, 每个重复(桶/260 L)60尾鱼。每天人工饱食投喂2次(7: 00和 18: 00), 在投喂结束后1h, 吸出残饵在70℃烘箱中烘干至恒重, 并称重, 每天记录投饵量。养殖周期 66d, 养殖期间记录每桶的死鱼数量并称重。实验条件: 水温, 25—26℃; 溶氧, >6.0 mg/L; pH, 6.8—7.6; 总氨氮量, <0.1 mg/L; 亚硝酸盐, <0.1 mg/L; 所有养殖桶均连续充气, 光照为自然光照。
养殖中期(35d)和养殖末期(66d)采集样品。取样前, 将鱼体饥饿24h, 统计每桶瓦氏黄颡鱼数量和重量, 每桶随机挑选10尾鱼, 用MS-222麻醉(1﹕ 10000; 上海康汀生物科技有限公司, 中国), 采集鱼体前肠和肝脏组织, 立即置于液氮, 待样本采集完毕后统一放于−80℃冰箱长期保存, 用于后续代谢基因的定量分析。实验末期每桶另取6尾黄颡鱼, 于−20℃冰箱保存, 用于后续体组成分析。
1.3 化学分析
采用AOAC (1995)[23]标准方法对饲料原料、饲料和鱼体成分进行分析; 采用全自动凯氏定氮仪(Kjeltec 2300, Sweden)测定粗蛋白含量(N×6.25); 采用索氏抽提法测定粗脂肪含量(Buchi 36680, Switzerland); 样品于电炉上炭化后, 马弗炉中(550℃)灼烧12h, 可得样品灰分含量。
1.4 RNA 提取
将黄颡鱼肝脏和前肠在液氮中磨成粉末, 然后放入含1 mL Trizol的无RNase离心管中, 分别加入氯仿、异丙醇、乙醇等试剂, 最后用灭菌水溶解沉淀, 具体操作过程参照李明珠[24]方法。RNA质量通过1.2%琼脂糖凝胶电泳检测, 浓度通过核酸定量仪(Nano Drop 2000 spectrophotometer, Thermo, USA)测定。将各组织样品的RNA调成1 μg后, 用PrimerScript®Reverse Transcriptase (TaKaRa, Japan)试剂盒将RNA反转成cDNA, 具体操作参照试剂盒说明书。
1.5 实时荧光定量PCR
采用SYBR®Green I嵌合荧光法, 使用SYBR®Premix Ex TaqTM(Perfect Real Time, TaKaRa, Japan)试剂在Thermal cycler (Mastercycler ep realplex, Eppendorf, German)仪器上进行, 反应条件为: 95℃ 2min, 1循环; 95℃变性15s, 59℃退火15s, 72℃延伸30s, 共计40个循环。溶解曲线: 95℃, 15s; 59℃, 15s, 95℃, 5min。以GAPDH和β-actin作为内参基因。在定量前, 验证内参基因、PEPT1和TOR2基因引物(表 2)的扩增效率。本实验PEPT1、TOR、GAPDH和β-actin基因的扩增效率分别为0.98、0.95、0.99和0.98, 随后以Lg(模板相对拷贝数)为横坐标, 以平均ΔCt(目的基因Ct值)−平均ΔCt(目的内参−内参基因Ct值后再平均)为纵坐标, 得到另外一条直线, 若其斜率绝对值小于0.1, 即说明目的基因与内参基因扩增效率一致, 用2–ΔΔCt进行基因表达分析[25]。分别对中期、末期瓦氏黄颡鱼PEPT1和TOR基因进行RT-PCR分析, 以水解鱼肉蛋白组为对照组, 对其他组的基因表达进行分析。
表 2 瓦氏黄颡鱼PEPT1和TOR2基因定量所需引物序列Table 2. Real-time quantitative PCR primers for PEPT1 and TOR2 genes基因Target gene 引物Primer (5′-3′) PEPT1 GCGTCATCCAAGAGTGCCC
ATCTGATTTGCTCCTCCGAGTOR2 GTGCAAGGACCTGACTCCAAGCC
TGATAGACTGGATGCGTATGATTGGGAPDH CATCCAAGGCGGCTGTCAAGCAAG
CCAAAGTTGTCGTTGAGGGCAAATGCβ-actin CACTGTGCCCATCTACGAG
CCATCTCCTGCTCGCAAGTC1.6 统计分析
生长指标计算如下:
成活率(SR, %)=100%×(末鱼数量/初鱼数量)
特定生长率(SGR, %/d)=100%×[(Ln终末重−Ln初始重)/实验天数]
结果采用SPSS17.0软件进行数据处理, 在单因素方差分析(ANOVA)数据达到显著(P<0.05)时, 进行Tukey’s多重比较, 数据以平均值±标准误(Mean±SEM)表示。
2. 结果
2.1 不同蛋白源对瓦氏黄颡鱼幼鱼不同生长阶段存活率和生长的影响
各处理组瓦氏黄颡鱼存活率(SR)无显著差异(P>0.05), 且SR均大于94%。在生长方面, 不同蛋白源显著影响瓦氏黄颡鱼生长性能(P<0.05)。在第35和第66天时, FM组瓦氏黄颡鱼SGR分别为4.86%/d和3.08%/d, SPC组瓦氏黄颡鱼SGR分别为3.94%/d和2.55%/d, SPC组瓦氏黄颡鱼SGR虽然低于FM组, 但显著高于FH组(1.13%/d和0.96%/d)和CAA组(1.69%/d和1.28%/d) (表 3)。
表 3 不同蛋白源对瓦氏黄颡鱼幼鱼不同生长阶段存活率和生长的影响(平均值±标准误, n=3)Table 3. Growth performance and survival of darkbarbel catfish (Pelteobagrus vachelli) fed diets at different growth stages (Mean±SEM, n=3)指标Index 饲料编号(蛋白源)Diet number (protein source) Diet 1 (FM) Diet 2 (SPC) Diet 3 (FH) Diet 4 (CAA) 初始体重IBW (g) 2.89±0.02 2.90±0.01 2.91±0.02 2.90±0.01 35d时体重35d-BW (g) 15.58±0.59a 11.54±0.26b 4.32±0.23c 5.28±0.25c 35d时特定生长率35d-SGR (%/d) 4.86±0.16a 3.94±0.24b 1.13±0.15d 1.69±0.13c 35d时存活率35d-SR (%) 97.49±0.03 98.74±0.04 100.00±0.03 100.00±0.03 66d时体重66d-BW (g) 22.22±0.15a 15.60±0.76b 5.48±0.25c 6.79±0.04c 66d时特定生长率66d-SGR (%/d) 3.08±0.18a 2.55±0.13b 0.96±0.09d 1.28±0.11c 66d时存活率66d-SR (%) 97.49±0.03 94.97±0.03 100.00±0.03 100.00±0.03 注: 表中数据为3个重复的平均值; 同一行相同右上角含有相同英文上标字母或无上标表示无显著差异(P≥0.05); 下同Note: Data are means of triplicates. Means in each bar sharing the same superscript letter or absence of superscripts are not significantly different determined by Tukey’s test (P≥0.05), the same applies below 2.2 不同蛋白源对瓦氏黄颡鱼幼鱼体组成的影响
不同蛋白源显著影响了瓦氏黄颡鱼体组成。SPC替代FM会降低鱼体粗蛋白含量, SPC组鱼体蛋白含量仅次于FM组, 其粗蛋白含量为13.28%。SPC组瓦氏黄颡鱼的粗脂肪含量显著高于FM组(P<0.05), 粗脂肪含量为 14.86%, SPC组瓦氏黄颡鱼灰分和水分含量显著低于其他组(P<0.05), 分别为6.80%和69.48% (表 4)。
表 4 不同蛋白源对瓦氏黄颡鱼幼鱼体组成的影响(平均值±标准误, n=3, 湿重%)Table 4. Body composition analysis of darkbarbel catfish (Pelteobagrus vachelli) fed diets at different growth stages (Mean±SEM, n=3, wet weight %)体组成Whole-body composition (%) 饲料编号(蛋白源)Diet number (protein source) Diet 1 (FM) Diet 2 (SPC) Diet 3 (FH) Diet 4 (CAA) 粗蛋白Crude protein 14.21±0.03a 13.28±0.01b 12.52±0.03c 11.44±0.07d 粗脂肪Crude lipid 12.36±0.12c 14.86±0.01a 10.73±0.03d 13.89±0.05ab 灰分Ash 9.06±0.03b 6.80±0.10d 9.59±0.12a 7.92±0.04c 水分Moisture 70.25±0.28bc 69.48±0.23c 74.05±0.05a 71.00±0.23b 2.3 不同蛋白源对瓦氏黄颡鱼幼鱼前肠PEPT1和肝脏TOR基因表达的影响
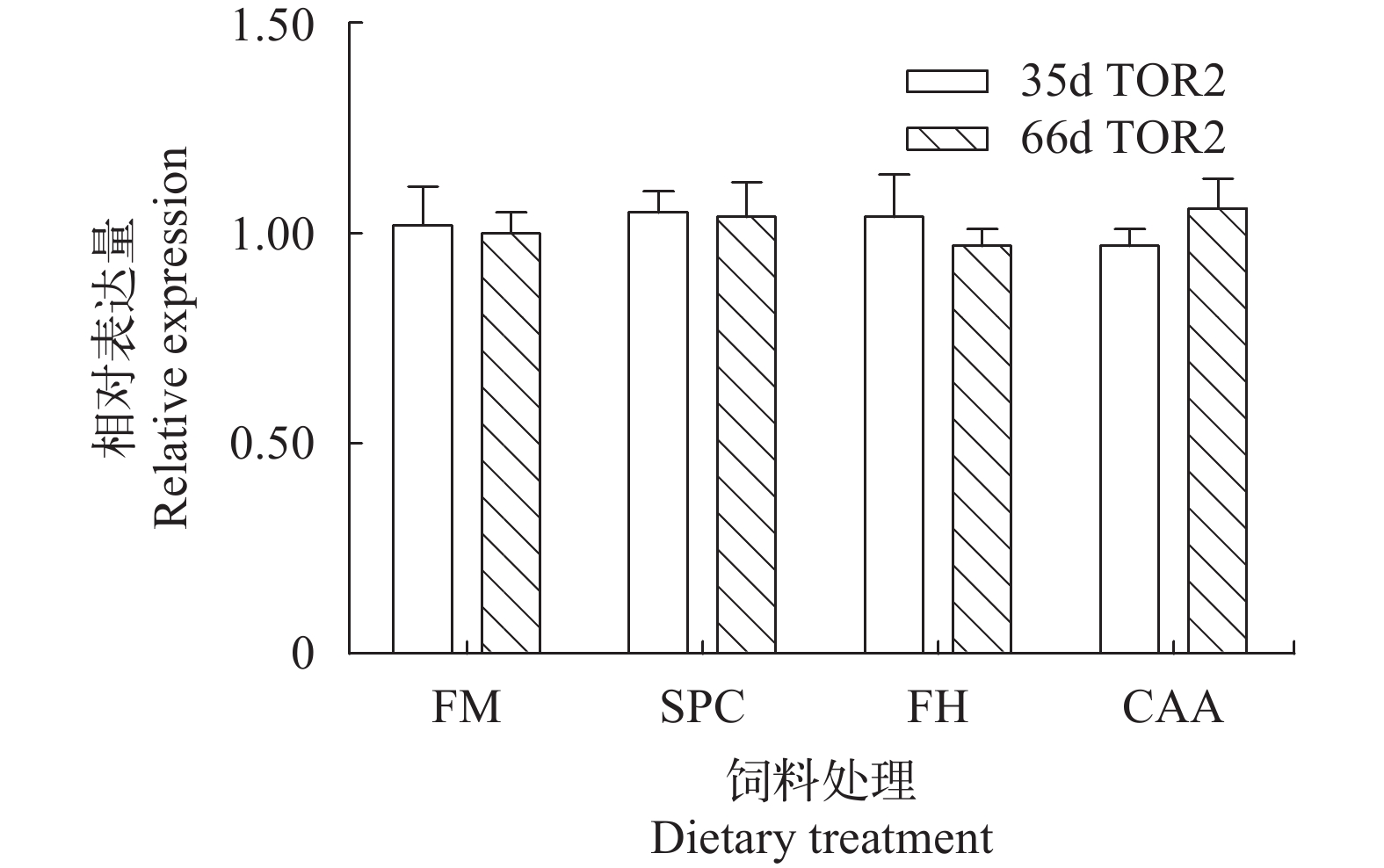
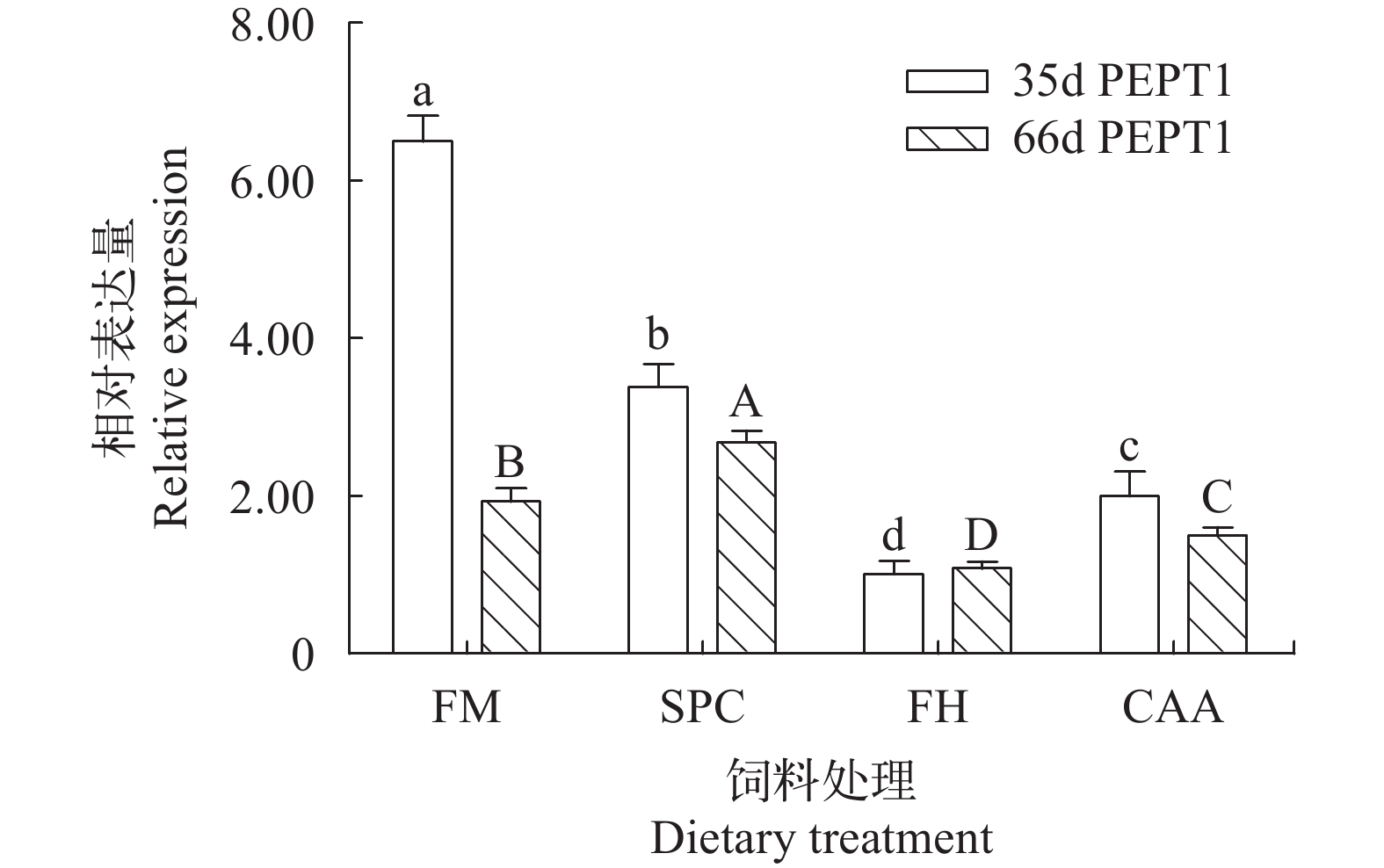
在第35和第66天时, 不同蛋白源显著影响了瓦氏黄颡鱼幼鱼前肠PEPT1的表达(P<0.05, 图 1)。在第35天时, FM组鱼体前肠PEPT1基因表达量显著高于其他处理组(P<0.05), SPC组鱼体前肠PEPT1表达量显著高于FH和CAA组(P<0.05)。在第66天时, SPC组鱼体前肠PEPT1表达量显著高于FM组(P<0.05)。在第35和第66天时, 不同蛋白源并未显著影响瓦氏黄颡鱼肝脏TOR基因的表达水平(P>0.05, 图 2)。
![]() 图 1 不同蛋白源对瓦氏黄颡鱼前肠PEPT1 mRNA表达的影响图中数值为平均值±标准误(n=3), 含有相同字母或没有上标代表差异不显著(P≥0.05); 下同Figure 1. Effects of different dietary protein sources on PEPT1 mRNA expressions in the foregut of darkbarbel catfishData are presented as means±SEM (n=3). Means in each bar sharing the same superscript letter or absence of superscripts are not significantly different determined by Tukey's test (P≥0.05), the same applies below
图 1 不同蛋白源对瓦氏黄颡鱼前肠PEPT1 mRNA表达的影响图中数值为平均值±标准误(n=3), 含有相同字母或没有上标代表差异不显著(P≥0.05); 下同Figure 1. Effects of different dietary protein sources on PEPT1 mRNA expressions in the foregut of darkbarbel catfishData are presented as means±SEM (n=3). Means in each bar sharing the same superscript letter or absence of superscripts are not significantly different determined by Tukey's test (P≥0.05), the same applies below![]() 图 2 不同蛋白源对瓦氏黄颡鱼肝脏TOR mRNA表达的影响Figure 2. Effects of different dietary protein sources on TOR mRNA expressions in the liver of darkbarbel catfish
图 2 不同蛋白源对瓦氏黄颡鱼肝脏TOR mRNA表达的影响Figure 2. Effects of different dietary protein sources on TOR mRNA expressions in the liver of darkbarbel catfish3. 讨论
3.1 不同蛋白源对瓦氏黄颡鱼幼鱼不同生长阶段存活率和生长的影响
本实验发现4种蛋白源对瓦氏黄颡鱼存活率无显著影响, 说明瓦氏黄颡鱼对养殖条件和实验饲料较为适应。在第35和第66天时, FM组实验鱼的生长性能达到最大, 显著高于其他各实验组; 通过比较瓦氏黄颡鱼SGR发现, SPC组虽然低于FM组, 但显著高于FH组和CAA组, 说明SPC是较理想的鱼粉替代源。SPC组的瓦氏黄颡鱼具有相对较好的生长性状, 这可能和SPC的特性相关, SPC蛋白含量高达65%—70%, 抗营养因子含量低, 同时SPC具有利用率高等特点[26], 因此, 使用SPC饲喂鱼体能使鱼体获得较好的生长性能。在大黄鱼[27]、真鲷(Pagrus major)[28]、虹鳟(Oncorhynchus mykiss)[29]中使用SPC进行投喂, 鱼体均具有较高的SGR。但是SPC缺乏鱼体生长所需的蛋氨酸, 因此本实验SPC的替代效果稍差于鱼粉, 这可能与SPC中缺乏蛋氨酸有关[30, 31]。
3.2 不同蛋白源对瓦氏黄颡鱼幼鱼体组成的影响
蛋白源会影响鱼体组成, 在本实验中鱼粉组瓦氏黄颡鱼的粗蛋白含量显著高于其他组。这可能与鱼粉具有较高的氨基酸平衡性有关[32—34], 鱼粉水解出的氨基酸更有利于鱼体蛋白沉积, 而这一点在其他实验中也得到类似的结果。周晖等[35]用酵母粉、脱脂豆粕和玉米蛋白粉分别替代10%的鱼粉, 饲喂体重30—38 g的军曹鱼(Rachycentron canadum)幼鱼5周, 结果发现FM组军曹鱼体蛋白含量最高。李学丽等[36]用2种豆粕部分替代鱼粉, 发现鱼粉组珍珠龙胆石斑鱼(♀Epinephelus fuscoguttatus×♂Epinephelus lanceolatus)幼鱼的全鱼体蛋白含量显著高于其他组。本实验同样发现, SPC组瓦氏黄颡鱼的体脂含量达到最大。与其他鱼粉替代源相比, SPC含有较少的抗营养因子, 虽然其氨基酸平衡性稍差于鱼粉, 但仍优于其他植物蛋白源, 有利于蛋白或糖类等转化为脂肪储存。SPC的这种特性在其他研究中也得到验证, Kissinger等[37]以SPC和其他蛋白源替代鱼粉饲喂长鳍(Seriola rivoliana) 9周, 发现其他组鱼体粗脂肪含量显著低于80%SPC组。
3.3 不同蛋白源对瓦氏黄颡鱼幼鱼前肠PEPT1和肝脏TOR基因表达的影响
不同蛋白源会显著影响鱼体蛋白质代谢相关基因的表达, 对虹鳟[38]饲喂含小肽、游离氨基酸和完整蛋白质的饲料会显著影响虹鳟肠道PEPT1的表达。在本实验中, 第35天和第66天时, FM组和SPC组黄颡鱼前肠PEPT1的表达高于其他组。相比于CAA、SPC和FM可被鱼体肠道蛋白酶水解为小肽, 可提高PEPT1 mRNA的表达, 促进鱼体对蛋白的吸收, 从而增加鱼体的蛋白沉积, 有利于鱼体生长, 这表明SPC是较好的FM替代源。在33d和66d时, FH组鱼体PEPT1 mRNA表达量最低, 在大菱鲆(Scophthalmus maximus L.) [39]上也发现类似结果。然而, 在大西洋鲑(Salmo salar L.)[40]、鲤(Cyprinus carpio L.) [41]和黄金鲈(Perca flavescens)[42]的相关研究中却得到不同的结果, 这很可能是由于鱼种、水解底物等不同所引起的。
PEPT1的表达还和生长阶段、能量水平等因素有关。在早期发育阶段, 鳜(Siniperca chuatsi)[43]肠道PEPT1表达量较高, 而随着鱼体体重的进一步增加, 肠道PEPT1的表达量逐渐下降并趋于稳定。在能量影响PEPT1的研究中, 高能量(13.17 kJ/kg)组肉鸡空肠和回肠 PEPT1表达量显著低于低能量组(12.34 kJ/kg)[44]。在本实验中, 在33d时FM组鱼体PEPT1表达量较高, 而66d时FM组鱼体PEPT1表达量出现一定程度下降。这可能是FM相比于CAA和FH, FM能量更高, PEPT1表达上升有利于黄颡鱼吸收能量物质, 用于生长, 而随着黄颡鱼体重进一步增加, PEPT1表达出现一定程度地下降。
综上所述, SPC的替代效果尽管差于FM, 但显著优于FH和CAA, 这可能是由于SPC上调前肠PEPT1的表达、促进鱼体对小肽的吸收所致, 表明SPC可作为一种鱼粉替代的优质蛋白源。
-
![]()
图 1 不同蛋白源对瓦氏黄颡鱼前肠PEPT1 mRNA表达的影响
图中数值为平均值±标准误(n=3), 含有相同字母或没有上标代表差异不显著(P≥0.05); 下同
Figure 1. Effects of different dietary protein sources on PEPT1 mRNA expressions in the foregut of darkbarbel catfish
Data are presented as means±SEM (n=3). Means in each bar sharing the same superscript letter or absence of superscripts are not significantly different determined by Tukey's test (P≥0.05), the same applies below
![]()
图 2 不同蛋白源对瓦氏黄颡鱼肝脏TOR mRNA表达的影响
Figure 2. Effects of different dietary protein sources on TOR mRNA expressions in the liver of darkbarbel catfish
表 1 实验饲料配方和化学成分(%干重)
Table 1 Formulation and chemical composition of the experimental diets (% dry matter)
原料Ingredient 饲料编号Diet number Diet 1 (FM) Diet 2 (SPC) Diet 3 (FH) Diet 4 (CAA) 鱼粉Fish meal1 48.65 0.00 0.00 0.00 大豆浓缩蛋白SPC1 0.00 48.32 0.00 0.00 晶体氨基酸CAA2 0.00 0.00 0.00 32.50 水解鱼肉蛋白FH1 0.00 0.00 33.69 0.00 小麦粉Wheat gluten1 25.00 25.00 25.00 25.00 麦麸Wheat bran1 13.00 13.00 13.00 13.00 鱼油Fish oil 0.27 4.50 4.41 4.50 豆油Soybean oil 1.81 1.17 1.81 1.81 卵磷脂Lecithin 2.00 2.00 2.00 2.00 矿物质预混料MP3 2.00 2.00 2.00 2.00 维生素预混料VP4 2.00 2.00 2.00 2.00 诱食剂Attractant5 0.50 0.50 0.50 0.50 抗氧化剂Antioxidant 0.05 0.05 0.05 0.05 微晶纤维素MC 4.72 1.46 15.54 16.64 成分分析Proximate composition (% dry matter)5 粗蛋白Crude protein (%) 39.20 39.13 39.30 39.17 粗脂肪Crude lipid (%) 8.60 8.97 9.03 8.79 灰分Ash (%) 6.35 7.33 9.11 10.21 注: 1鱼粉: 粗蛋白占干重66.80%, 粗脂肪占干重8.70%; 大豆浓缩蛋白: 粗蛋白占干重67.26%, 粗脂肪占干重1.31%; 水解鱼肉蛋白: 粗蛋白占干重84.99%, 粗脂肪占干重3.19%; 小麦粉: 粗蛋白占干重16.10%, 粗脂肪占干重1.36%; 麦麸: 粗蛋白占干重19.04%, 粗脂肪占干重2.73%; 2晶体氨基酸(g/kg diet): 精氨酸, 3.42 g; 组氨酸, 0.26 g; 异亮氨酸, 2.52 g; 亮氨酸, 4.36 g; 赖氨酸, 4.26 g; 蛋氨酸, 1.57 g; 苯丙氨酸, 2.29 g; 苏氨酸, 2.41 g; 缬氨酸, 3.06 g; 丙氨酸, 3.67 g; 天冬氨酸, 5.65 g; 胱氨酸, 0.67 g; 谷氨酸, 8.48 g; 甘氨酸, 3.36 g; 丝氨酸, 2.41 g; 酪氨酸, 1.46 g; 脯氨酸, 2.11 g; 3矿物质预混料(mg or g/kg diet): 氟化钠, 2 mg; 碘化钾, 0.8 mg; 氯化钴(1%), 50 mg; 硫酸铜, 10 mg; 硫酸铁, 80 mg; 硫酸锌, 50 mg; 硫酸锰, 60 mg; 硫酸镁, 1200 mg; 磷酸二氢钙, 3000 mg; 氯化钠, 100 mg; 沸石粉, 15.448 g; 4维生素预混料(mg or g/kg diet): 硫胺素, 25 mg; 维生素B2, 45 mg; 盐酸吡哆醇, 20 mg; 维生素B12, 0.1 mg; 维生素K3, 10 mg; 肌醇, 800 mg; 泛酸, 60 mg; 烟酸, 200 mg; 叶酸, 20 mg; 生物素, 1.20 mg; 维生素A, 32 mg; 维生素D, 5 mg; 维生素E, 120 mg; 维生素C, 2000 mg; 胆碱, 2000 mg; 乙氧基喹啉, 150 mg; 微晶纤维素, 14.52 g; 5诱食剂: 甘氨酸和甜菜碱Note: 1Fish meal: crude protein 66.80% dry matter, crude lipid 8.70% dry matter; SPC: crude protein 67.26% dry matter, crude lipid 1.31% dry matter; Fish hydrolysate: crude protein 84.99% dry matter, crude lipid 3.19% dry matter; Wheat gluten: crude protein 16.10% dry matter, crude lipid 1.36% dry matter; Wheat bran: crude protein 19.04% dry matter, crude lipid 2.73% dry matter; 2Crystalline amino acids (g/kg diet): Arginine, 3.42 g; Histidine, 0.26 g; Isoleucine, 2.52 g; Leucine, 4.36 g; Lysine, 4.26 g; Methionine, 1.57 g; Phenylalanine, 2.29 g; Threonine, 2.41 g; Valine, 3.06 g; Alanine, 3.67 g; Aspartic acid, 5.65 g; Cystine, 0.67 g; Glutamic acid, 8.48 g; Glycine, 3.36 g; Serine, 2.41 g; Tyrosine, 1.46 g; Proline, 2.11 g; 3Mineral premix (mg or g/kg diet): NaF, 2 mg; KI, 0.8 mg; CoCl2·6H2O(1%), 50 mg; CuSO4·5H2O, 10 mg; FeSO4·H2O, 80 mg; ZnSO4·H2O, 50 mg; MnSO4·H2O, 60 mg; MgSO4·7H2O, 1200 mg; Ca(H2PO4)2·H2O, 3000 mg; NaCl, 100 mg; Zoelite, 15.448 g; 4Vitamin premix (mg or g/kg diet): thiamin, 25 mg; riboflavin, 45 mg; pyridoxine-HCl, 20 mg; vitamin B12, 0.1 mg; vitamin K3, 10 mg; inositol, 800 mg; pantothenic acid, 60 mg; niacin acid, 200 mg; folic acid, 20 mg; biotin, 1.20 mg; retinol acetate, 32 mg; cholecalciferol, 5 mg; alpha-tocopherol, 120 mg; ascorbic acid, 2000 mg; choline chloride, 2000 mg; ethoxyquin, 150 mg; microcrystalline cellulose, 14.52 g; 5Attractant: glycine and betaine  下载: 导出CSV
下载: 导出CSV
表 2 瓦氏黄颡鱼PEPT1和TOR2基因定量所需引物序列
Table 2 Real-time quantitative PCR primers for PEPT1 and TOR2 genes
基因Target gene 引物Primer (5′-3′) PEPT1 GCGTCATCCAAGAGTGCCC
ATCTGATTTGCTCCTCCGAGTOR2 GTGCAAGGACCTGACTCCAAGCC
TGATAGACTGGATGCGTATGATTGGGAPDH CATCCAAGGCGGCTGTCAAGCAAG
CCAAAGTTGTCGTTGAGGGCAAATGCβ-actin CACTGTGCCCATCTACGAG
CCATCTCCTGCTCGCAAGTC
下载: 导出CSV
表 3 不同蛋白源对瓦氏黄颡鱼幼鱼不同生长阶段存活率和生长的影响(平均值±标准误, n=3)
Table 3 Growth performance and survival of darkbarbel catfish (Pelteobagrus vachelli) fed diets at different growth stages (Mean±SEM, n=3)
指标Index 饲料编号(蛋白源)Diet number (protein source) Diet 1 (FM) Diet 2 (SPC) Diet 3 (FH) Diet 4 (CAA) 初始体重IBW (g) 2.89±0.02 2.90±0.01 2.91±0.02 2.90±0.01 35d时体重35d-BW (g) 15.58±0.59a 11.54±0.26b 4.32±0.23c 5.28±0.25c 35d时特定生长率35d-SGR (%/d) 4.86±0.16a 3.94±0.24b 1.13±0.15d 1.69±0.13c 35d时存活率35d-SR (%) 97.49±0.03 98.74±0.04 100.00±0.03 100.00±0.03 66d时体重66d-BW (g) 22.22±0.15a 15.60±0.76b 5.48±0.25c 6.79±0.04c 66d时特定生长率66d-SGR (%/d) 3.08±0.18a 2.55±0.13b 0.96±0.09d 1.28±0.11c 66d时存活率66d-SR (%) 97.49±0.03 94.97±0.03 100.00±0.03 100.00±0.03 注: 表中数据为3个重复的平均值; 同一行相同右上角含有相同英文上标字母或无上标表示无显著差异(P≥0.05); 下同Note: Data are means of triplicates. Means in each bar sharing the same superscript letter or absence of superscripts are not significantly different determined by Tukey’s test (P≥0.05), the same applies below
下载: 导出CSV
表 4 不同蛋白源对瓦氏黄颡鱼幼鱼体组成的影响(平均值±标准误, n=3, 湿重%)
Table 4 Body composition analysis of darkbarbel catfish (Pelteobagrus vachelli) fed diets at different growth stages (Mean±SEM, n=3, wet weight %)
体组成Whole-body composition (%) 饲料编号(蛋白源)Diet number (protein source) Diet 1 (FM) Diet 2 (SPC) Diet 3 (FH) Diet 4 (CAA) 粗蛋白Crude protein 14.21±0.03a 13.28±0.01b 12.52±0.03c 11.44±0.07d 粗脂肪Crude lipid 12.36±0.12c 14.86±0.01a 10.73±0.03d 13.89±0.05ab 灰分Ash 9.06±0.03b 6.80±0.10d 9.59±0.12a 7.92±0.04c 水分Moisture 70.25±0.28bc 69.48±0.23c 74.05±0.05a 71.00±0.23b
下载: 导出CSV
-
[1] Kaushik S J, Cravedi J P, Lalles J P, et al. Partial or total replacement of fish meal by soybean protein on growth, protein utilization, potential estrogenic or antigenic effects, cholesterolemia and flesh quality in rainbow trout, Oncorhynchus mykiss [J]. Aquaculture, 1995, 133(3): 257—274
[2] García V, Celada J D, González R, et al. Response of juvenile tench (Tinca tinca L.) fed practical diets with different protein contents and substitution levels of fish meal by soybean meal [J]. Aquaculture Research, 2015, 46(1): 28—38 doi: 10.1111/are.2014.46.issue-1
[3] Kaushik S J, Coves D, Dutto G, et al. Almost total replacement of fish meal by plant protein sources in the diet of a marine teleost, the European seabass, Dicentrarchus labrax [J]. Aquaculture, 2004, 230(1—4): 391—404 doi: 10.1016/S0044-8486(03)00422-8
[4] Tacon A G, Metian M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects [J]. Aquaculture, 2008, 285(1): 146—158
[5] 艾庆辉, 谢小军. 水生动物对植物蛋白源利用的研究进展. 中国海洋大学学报(自然科学版), 2005, 6: 51—57 Ai Q H, Xie X J. Advance in utilization of plant proteins by aquatic animals [J]. Journal of Ocean University of China, 2005, 6: 51—57
[6] Berge G M, Grisdale-Helland B, Helland S J. Soy protein concentrate in diets for Atlantic halibut (Hippoglossus hippoglossus) [J]. Aquaculture, 1999, 178(1): 139—148
[7] Deng J M, Mai K S, Ai Q H, et al. Effects of replacing fish meal with soy protein concentrate on feed intake and growth of juvenile Japanese flounder, Paralichthys olivaceus [J]. Aquaculture, 2006, 258(1): 503—513
[8] Hien T T T, Phu T M, Tu T L C, et al. Effects of replacing fish meal with soya protein concentrate on growth, feed efficiency and digestibility in diets for snakehead, Channa striata [J]. Aquaculture Research, 2017, 48(6): 3174—3181 doi: 10.1111/are.2017.48.issue-6
[9] 宋志东, 王际英, 李培玉, 等. 酶解大豆蛋白替代鱼粉对星斑川鲽幼鱼生长、血液生化和体组成的影响. 水生生物学报, 2016, 40(1): 165—172 Song Z D, Wang J Y, Li P Y, et al. Effects of replacement fishmeal with soy protein hydrolysates on growth, blood biochemistry, body composition of juvenile starry flounder Platichthys stellatus Pallas [J]. Acta Hydrobiologica Sinica, 2016, 40(1): 165—172
[10] Khosravi S, Bui H T D, Rahimnejad S, et al. Dietary supplementation of marine protein hydrolysates in fish-meal based diets for red sea bream (Pagrus major) and olive flounder (Paralichthys olivaceus) [J]. Aquaculture, 2015, 435: 371—376 doi: 10.1016/j.aquaculture.2014.10.019
[11] Aksnes A, Hope B, Høstmark Ø, et al. Inclusion of size fractionated fish hydrolysate in high plant protein diets for Atlantic cod, Gadus morhua [J]. Aquaculture, 2006, 261(3): 1102—1110 doi: 10.1016/j.aquaculture.2006.07.038
[12] Cai Z N, Li W J, Mai K S, et al. Effects of dietary size-fractionated fish hydrolysates on growth, activities of digestive enzymes and aminotransferases and expression of some protein metabolism related genes in large yellow croaker (Larimichthys crocea) larvae [J]. Aquaculture, 2015, 440: 40—47 doi: 10.1016/j.aquaculture.2015.01.026
[13] 刘经纬, 麦康森, 徐玮, 等. 谷氨酰胺对半滑舌鳎稚鱼非特异性免疫相关酶活力和低氧应激后HIF-1a表达的影响. 水生生物学报, 2016, 40(4): 736—743 Liu J W, Mai K S, Xu W, et al. Effects of dietary glutamine on activities of non-specific immune related enzymes and HIF-1a expression after hypoxia of half-smooth tongue sole post larvae [J]. Acta Hydrobiologica Sinica, 2016, 40(4): 736—743
[14] 王震, 徐玮, 麦康森, 等. 饲料缬氨酸水平对军曹鱼鱼体脂肪含量、血浆生化指标和肝脏脂肪代谢基因表达的影响. 水生生物学报, 2016, 40(4): 744—751 Wang Z, Xu W, Mai K S, et al. The effects of valine level on plasma biochemical indexes, lipid content and gene expression involved in lipid metabolism in cobia (Rachycentron canadum) [J]. Acta Hydrobiologica Sinica, 2016, 40(4): 744—751
[15] 张珊, 艾庆辉, 麦康森, 等. 晶体氨基酸替代鱼粉蛋白对半滑舌鳎稚鱼消化酶和代谢酶活力的影响. 水生生物学报, 2014, 38(5): 801—808 doi: 10.7541/2014.121 Zhang S, Ai Q H, Mai K S, et al. Effects of fish meal replacement with crystalline amino acid on digestive and metabolic enzymes of tongue sole (Cynoglossus semilaevis) larae [J]. Acta Hydrobiologica Sinica, 2014, 38(5): 801—808 doi: 10.7541/2014.121
[16] Terova G, Robaina L, Izquierdo M, et al. PepT1 mRNA expression levels in sea bream (Sparus aurata) fed different plant protein sources [J]. Springerplus, 2013, 2(1): 1—14 doi: 10.1186/2193-1801-2-1
[17] Spanier B. Transcriptional and functional regulation of the intestinal peptide transporter PEPT1 [J]. Journal of Physiology, 2014, 592(5): 871—879 doi: 10.1113/jphysiol.2013.258889
[18] Dumas A, Lange C F, France J, et al. Quantitative description of body composition and rates of nutrient deposition in rainbow trout (Oncorhynchus mykiss) [J]. Aquaculture, 2007, 273(1): 165—181 doi: 10.1016/j.aquaculture.2007.09.026
[19] Liu Z, Barrett E J. Human protein metabolism: its measurement and regulation [J]. American Journal of Physiology Endocrinology & Metabolism, 2002, 283(6): E1105—E1112
[20] Song S, Hooiveld G J, Li M, et al. Dietary soy and meat proteins induce distinct physiological and gene expression changes in rats [J]. Scientific Reports, 2016, 6: 20036 doi: 10.1038/srep20036
[21] Spanier B, Rohm F. Proton coupled oligopeptide transporter 1 (PepT1) function, regulation, and influence on the intestinal homeostasis [J]. Comprehensive Physiology, 2018, 8(2): 843—869
[22] Zheng K K, Zhu X M, Han D, et al. Effects of dietary lipid levels on growth, survival and lipid metabolism during early ontogeny of Pelteobagrus vachelli larvae [J]. Aquaculture, 2010, 299(1): 121—127
[23] Cuniff P E. Official Methods of Analysis of AOAC International: Agricultural Chemicals; Contaminants; Drugs [M]. AOAC International. 1995, 1899
[24] 李明珠. 皱纹盘鲍(Haliotis discus hannai Ino.)Δ6脂肪酸去饱和酶(Δ6FAD)基因克隆以及不同脂肪源对皱纹盘鲍稚鲍生长、脂肪酸组成和Δ6FAD表达的影响. 中国海洋大学. 2010 Li M Z. Cloning of Δ6 fatty acid desaturase (Δ6FAD) and the effect of different dietary lipids on growth, fatty acid composition and Δ6FAD mRNA relative expression level of juvenile abalone (Haliotis discus hannai Ino.) [D]. Thesis for master of Science. Ocean University of China, Qingdao. 2010
[25] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔC t method [J]. Methods, 2001, 25(4): 402—408 doi: 10.1006/meth.2001.1262
[26] Aragao C, Lec C, Dias J, et al. Soy protein concentrate as a protein source for Senegalese sole (Solea senegalensis Kaup 1858) diets: effects on growth and amino acid metabolism of postlarvae [J]. Aquaculture Research, 2015, 34(15): 1443—1452
[27] Wang P, Zhu J Q, Feng J, et al. Effects of dietary soy protein concentrate meal on growth, immunity, enzyme activity and protein metabolism in relation to gene expression in large yellow croaker Larimichthys crocea [J]. Aquaculture, 2017, 477: 15—22 doi: 10.1016/j.aquaculture.2017.04.030
[28] Takagi S T, Shimeno S, Hosokawa H, et al. Effect of lysine and methionine supplementation to a soy protein concentrate diet for red sea bream Pagrus major [J]. Fisheries Science, 2010, 67(6): 1088—1096
[29] Stickney R R, Hardy R W, Koch K, et al. The effects of substituting selected oilseed protein concentrates for fish meal in rainbow trout Oncorhynchus mykiss diets [J]. Journal of the World Aquaculture Society, 2010, 27(1): 57—63
[30] Bañuelos-Vargas I, López L M, Pérez-Jiménez A, et al. Effect of fishmeal replacement by soy protein concentrate with taurine supplementation on hepatic intermediary metabolism and antioxidant status of totoaba juveniles (Totoaba macdonaldi) [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2014, 170: 18—25 doi: 10.1016/j.cbpb.2014.01.003
[31] Kokou F, Sarropoulou E, Cotou E, et al. Effects of graded dietary levels of soy protein concentrate supplemented with methionine and phosphate on the immune and antioxidant responses of gilthead sea bream (Sparus aurata L.) [J]. Fish & Shellfish Immunology, 2017, 64: 111—121
[32] 艾庆辉, 谢小军. 南方鲇的营养学研究: 饲料中大豆蛋白水平对消化率及摄食率的影响. 水生生物学报, 2002, 26(3): 215—220 doi: 10.3321/j.issn:1000-3207.2002.03.002 Ai Q H, Xie X J. The nutrition of silurus meridionalis: effects of different levels of dietary soybean protein on digestibility and feeding rate [J]. Acta Hydrobiologica Sinica, 2002, 26(3): 215—220 doi: 10.3321/j.issn:1000-3207.2002.03.002
[33] 何娇娇, 王萍, 冯建, 等. 玉米蛋白粉替代鱼粉对大黄鱼生长、血清生化指标及肝脏组织学的影响. 水生生物学报, 2017, 41(3): 506—515 He J J, Wang P, Feng J, et al. Effects of replacing fish meal with corn gluten meal on the growth, serum biochemical indices and liver histology of large yellow croaker Larimichthys crocea [J]. Acta Hydrobiologica Sinica, 2017, 41(3): 506—515
[34] 孙瑞健, 张文兵, 徐玮, 等. 饲料蛋白质水平与投喂频率对大黄鱼生长、体组成及蛋白质代谢的影响. 水生生物学报, 2013, 37(2): 281—289 Sun R J, Zhang W B, Xu W, et al. Effects of dietary protein level and feeding frequency on the growth performance, body composition and protein metabolism of juvenile large yellow croaker Larimichthys crocea R. [J]. Acta Hydrobiologica Sinica, 2013, 37(2): 281—289
[35] 周晖, 陈刚, 林小涛. 三种蛋白源部分替代鱼粉对军曹鱼幼鱼生长和体成分的影响. 水产科学, 2012, 31(6): 311—315 doi: 10.3969/j.issn.1003-1111.2012.06.001 Zhou H, Chen G, Lin X T. Effects of partial replacement of fish meal by three types of protein sources on growth performance and approximate compositions of juvenile cobia (Rachycentron canadum) [J]. Fisheries Science, 2012, 31(6): 311—315 doi: 10.3969/j.issn.1003-1111.2012.06.001
[36] 李学丽, 王际英, 宋志东, 等. 酶解豆粕替代鱼粉对珍珠龙胆石斑鱼幼鱼生长和主要代谢酶活力的影响. 海洋渔业, 2017, 39(5): 529—538 doi: 10.3969/j.issn.1004-2490.2017.05.006 Li X L, Wang J Y, Song Z D, et al. Effects of fishmeal replacement by hydrolyzed soybean meal on growth, body composition, digestive and metabolic enzyme activities of juvenile Epinephelus fuscoguttatus×Epinephelus lanceolatus [J]. Ocean Fishery, 2017, 39(5): 529—538 doi: 10.3969/j.issn.1004-2490.2017.05.006
[37] Kissinger K R, García-Ortega A, Trushenski J T. Partial fish meal replacement by soy protein concentrate, squid and algal meals in low fish-oil diets containing Schizochytrium limacinum for longfin yellowtail Seriola rivoliana [J]. Aquaculture, 2016, 452: 37—44 doi: 10.1016/j.aquaculture.2015.10.022
[38] Ostaszewska T, Kamaszewski M, Grochowski P, et al. The effect of peptide absorption on PepT1 gene expression and digestive system hormones in rainbow trout (Oncorhynchus mykiss) [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2010, 155(1): 107—114
[39] Wei Y L, Liang M Q, Mu Y, et al. The effect of ultrafiltered fish protein hydrolysate level on growth performance, protein digestibility and mRNA expression of PepT1 in juvenile turbot (Scophthalmus maximus L.) [J]. Aquaculture Nutrition, 2016, 22(5): 1006—1017 doi: 10.1111/anu.2016.22.issue-5
[40] Bakke S, Jordal A, Gómez-Requeni P, et al. Dietary protein hydrolysates and free amino acids affect the spatial expression of peptide transporter PepT1 in the digestive tract of Atlantic cod (Gadus morhua) [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2010, 156(1): 48—55 doi: 10.1016/j.cbpb.2010.02.002
[41] Ostaszewska T, Dabrowski K, Kamaszewski M, et al. The effect of plant protein-based diet supplemented with dipeptide or free amino acids on digestive tract morphology and PepT1 and PepT2 expressions in common carp (Cyprinus carpio L.) [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2010, 157(2): 158—169
[42] Ostaszewska T, Dabrowski K, Kamaszewski M, et al. The effect of dipeptide, Lys-Gly, supplemented diets on digestive tract histology in juvenile yellow perch (Perca flavescens) [J]. Aquaculture Nutrition, 2013, 19(1): 100—109 doi: 10.1111/j.1365-2095.2012.00948.x
[43] 刘知行, 张健东, 刘小燕, 等. 鳜小肽转运载体PEPT1基因分子特征及其表达研究. 水生生物学报, 2014, 38(3): 556—562 Liu Z X, Zhang J D, Liu X Y, et al. Molecular characterization and expression research of oligopeptide transporter PEPT1 in Siniperca chuatsi [J]. Acta Hydrobiologica Sinica, 2014, 38(3): 556—562
[44] 刘醒醒. 能量和粗蛋白质水平对肉仔鸡生产性能、养分消化和相关基因表达量的影响. 甘肃农业大学. 2016 Liu X X. The effect of dietary crude protein and energy level on performance, nutrient utilization and related genes in broiler [D]. Thesis for master of Science. Gansu Agricultural University, Lanzhou. 2016
-
期刊类型引用(1)
1. Heng ZHANG, Guoliang RUAN, Kun GUO, Wenhao FAN, Weiyou ZHENG. Study on Early Propagation Technology of Yellow Catfish (Pelteobagrus fulvidraco). Agricultural Biotechnology. 2020(01): 63-66+69 .  必应学术
必应学术
其他类型引用(0)
计量
- 文章访问数: 2467
- HTML全文浏览量: 709
- PDF下载量: 112
- 被引次数: 1